Changes in a Lady Beetle Community Following the Establishment of Three Alien Species
Andrei Alyokhin and Gary Sewell
Department of Biological Sciences, University of Maine, 5722 Deering Hall, Orono, ME 04469
This peer-refereed invited review article has been first published by Biological Invasions (Volume 6, Pages 463-471). The original publication is available at www.springerlink.com. Reproduced with permission of the publisher. Please send me an e-mail if you want a PDF reprint of this article in its original journal format.
ABSTRACT
A number of recent studies indicated that establishment of exotic lady beetles (Coleoptera: Coccinellidae) may have adverse affects on native lady beetle species. In the present study, we analyzed changes in coccinellid community inhabiting potato crops in northern Maine over the past 31 years. Prior to 1980, lady beetle communities were comprised almost exclusively of the two native species, Coccinella transversoguttata Brown and Hippodamia tredecimpunctata (Say). Starting 1980, an exotic species Coccinella septempunctata L. became permanently established in potato crops and quickly started to dominate lady beetle community. Two other exotic species, Harmonia axyridis (Pallas), and Propylea quatordecimpunctata (L.) became prominent members of the lady beetle community in 1995 and 1996. Invasion by exotic species was followed by a significant decline in the abundance of C. transversoguttata and H. tredecimpunctata, and a significant increase in the overall diversity of lady beetle community. The abundance of aphid prey was substantially reduced after the establishment of H. axyridis. The observed trends demonstrate the profound effects that exotic natural enemies may have on target and non-target native species, and highlight the importance of their thorough evaluation before initiating biological control programs.
Key words: biological invasion, biological control, Coccinellidae, exotic species, faunal change, interspecific interactions, natural enemies
Introduction
Invasion by exotic species has become one of the major environmental problems in a variety of ecosystems all over the world. Expanding human transport and commerce have led to a dramatic increase in the geographic scope, frequency, and number of exotic introductions (Mack et al. 2000). Furthermore, numerous species have been deliberately introduced into new locations for biological control of economically important pests (De Bach and Rosen 1991). Historically, most scientists and the general public viewed biological control as an efficient, low-cost, and environmentally safe approach to pest management (De Bach and Rosen 1991). However, serious concerns over negative impacts of natural enemy introductions have been recently raised by a number of prominent ecologists and conservation biologists (see reviews in Howarth 1991; Simberloff and Stiling 1996; Lockwood 1993, 1996; Hopper 1998). Therefore, a better understanding of natural enemy fate following introduction and its impacts on the new environment is essential for advancing theory and practice of biological control.
Lady beetles (Coccinellidae) are one of the most important groups that prey on pest insects, particularly on aphids and other Homoptera. Spectacular success of the introduced Vedalia beetle, Rodolia cardinalis (Mulsant) in controlling cottony cushion scale in California in late 19th century has ever since been a textbook example illustrating accomplishments of biological control (De Bach and Rosen 1991). Overall, a total of 179 lady beetle species have been deliberately or inadvertently introduced into the U.S. Of those, 26 species became established, often contributing to the suppression of insect pests (Gordon 1985). Unfortunately, a number of recent studies indicated that establishment of exotic lady beetles may have adverse effects on native lady beetle species. Increased competition for food, intraguild predation, and the combination of those two factors were suggested as possible explanations to the observed negative interactions among the species (Hironori and Katsuhiro 1991; Elliott et al. 1996; Obrycki et al. 1998; Evans 1991, 2000; Kajita et al. 2000).
Coccinella septempunctata L., Harmonia axyridis (Pallas), and Propylea quatordecimpunctata (L.) are three species of exotic lady beetles that are currently very common in eastern North America (Day et al. 1994; Wheeler and Hoebeke 1995; Hoebeke and Wheeler 1996; Ellis et al. 1999). All three of them are Old World in origin. C. septempunctata and P. quatordecimpunctata are widespread Palearctic species (Gordon 1985; Hoebeke and Wheeler 1996), while H. axyridis occurs in southern Siberia, Manchuria, China, Taiwan, Korea, Japan, Ryukyu Islands, and Bonin Islands (Gordon 1985; Chapin and Brou 1991), The exact mode of their introduction to North America is unknown. At one time or another, all three species have been deliberately released for biological control of pest aphids (Shands et al. 1972; Gordon 1985; Schaefer et al. 1987; Day et al. 1994). However, subsequent surveys at or near release sites usually failed to detect the establishment of these species. On the opposite, most of them were first recovered in the vicinity of seaports (Day et al. 1994). Therefore, it is generally assumed that their current populations in North America resulted from inadvertent introductions with ship cargoes (Day et al. 1994; Wheeler and Hoebeke 1995).
Based on an extensive literature review, Wheeler and Hoebeke (1995) suggested that invasion by C. septempunctata, a polyphagous lady beetle of paleoarctic origin, might have been responsible for the observed dramatic decline of the native lady beetle Coccinella novemnotata Herbst. Similarly, Elliott et al. (1996) observed 20-32 fold decrease in the abundance of native C. transversoguttata Brown and Adalia bipunctata (L.) after C. septempunctata establishment in alfalfa, corn, and small grain fields in eastern South Dakota. In turn, C. septempunctata has been recently replaced by H. axyridis (Pallas), in West Virginia apple orchards (Brown and Miller 1998). Clearly, invasion by exotic species is causing profound modifications in lady beetle communities, and a better understanding of such changes has important theoretical and practical implications.
Lady beetle populations commonly exhibit high temporal variability (Kieckhefer and Elliott 1990; Evans and Youssef 1992). Therefore, long-term studies conducted at the same location are essential to reliably delineate trends and patterns in ecological processes taking place in their populations (Elliott et al. 1996). Elliott et al. (1996) and Brown and Miller (1998) obtained extremely valuable information on the impacts of exotic lady beetles using 13 and 16-year data sets collected in South Dakota and West Virginia, respectively. In the present study, we further expanded this line of research by analyzing changes in coccinellid community inhabiting potato crops in northern Maine over the past 31 years.
Materials and Methods
Data used in the present study have been collected from the untreated control plots set up as a part of insecticide trials at the Aroostook Reasearch Farm, Presque Isle, Maine. The trials were conducted every year between 1971 and 2001, except for 1976 and 1979. Experimental protocols were very similar each year of investigations. Approximately 17.7 m long and 4 row wide experimental plots were planted to certified seed potato tubers that were cut into 75-80 g pieces prior to planting. The distance between the rows was ~90 cm, and seed spacing was ~35 cm. Approximately 1.8 m was left between the plots within each block, and blocks were spaced at ~3 m. All plots were fertilized with NPK fertilizer at planting and sprayed weekly with fungicides to control fungal diseases of potatoes. Between 5 and 30 plots were planted each year, depending on the number of chemicals tested. Potatoes were grown in a two-year rotation with small grains.
Twenty-five plants were randomly selected at weekly intervals from the middle two rows of each plot and visually examined for the presence of lady beetles. The number of adult Hippodamia tredecimpunctata (Say), Coccinella transversoguttata Brown (two native species), as well as the number of exotic C. septempunctata L., H. axyridis (Pallas), and P. quatordecimpunctata (L.) present on each selected plant was recorded. Together, these five species usually accounted for 100% of all lady beetles present at the potato plots. Lady beetles belonging to other species were found sporadically during only 3 years of the 31-year study. Those were recorded as ‘other’ without attempting to identify them to a species. Additionally, the number of potato-colonizing aphids (potato aphid, Macrosiphum euphorbiae (Thomas), buckthorn aphid, Aphis nasturtii Kaltenbach, and green peach aphid, Myzus persicae (Sulzer)) infesting one top, one middle, and one bottom leaf of each selected plant were counted at weekly intervals throughout the growing season. For operational reasons, the number of plants sampled at each plot was reduced to 20 starting 1995.
For each year of the study, mean seasonal insect density was calculated separately for each replication. Densities were expressed on per plant (for lady beetles) or per leaf (for aphids) basis. Because there is no evidence that any aphid species is a preferred prey for ladybeetles, all three of them were pooled together. The seasonal means were used for further statistical analysis.
Relative abundance of different lady beetle species at different years of the study was compared using Chi-square test (PROC FREQ, SAS Institute 1999). Species diversity was characterized using the Berger-Parker species dominance index (d) and the Shannon diversity index (H’) (Magurran 1988). Differences in species diversity among years, as well as differences in total abundances of lady beetle beetles and their aphid prey were analyzed by ANOVA (PROC GLM, SAS Institute 1999) (Magurran 1988; Wright and DeVries 2000). Data normality was checked before the analyses using the Wilk-Shapiro test (P=0.01) (PROC UNIVARIATE, SAS Institute 1999). Because of its non-normal distribution, the data were then transformed using rank transformations (Conover and Iman 1981) or arcsine transformations (Zar 1999). Means and standard errors presented in this article were calculated from the non-transformed data only. ANOVA was followed by orthogonal multiple contrasts that were designed a priori (Zar 1999) to test the differences between various time periods. Those were: (1) time period before 1980 when lady beetle complex consisted almost exclusively of native beetles vs. time period starting 1980 when exotic beetles became well established within experimental plots, (2) time period between 1980-1995 when C. septempunctata was the major exotic species vs. time period starting 1995 when H. axyridis and P. quatordecimpunctata also became prominent part of the community. Mutual interactions of different lady beetle species was analyzed using Spearman’s rank correlations (PROC CORR, SAS Institute 1999), and the impact of those species on the abundance of aphid prey was analyzed by multiple regression (PROC REG, SAS Institute 1999).
Additional information on lady beetle populations in the untreated control plots for the period between 1955 and 1964 was obtained from the archive maintained at the Aroostook Experiment Farm. Unfortunately, those records contained only information on relative abundance of different species, and not on their population densities. Also, no data were available for the period between 1964 and 1971. Therefore, only data collected starting 1971 were included in the statistical analysis.
Results
The relative abundance of different lady beetle species varied substantially among the years (Figs. 1 and 2), with the observed difference being highly significant (χ2= 452.8, df=28, P<0.0001). Prior to 1980, lady beetle communities were comprised almost exclusively of the two native species, H. tredecimpunctata and C. transversoguttata. Despite extensive releases of laboratory-reared eggs and larvae conducted in 1964-69 (Shands et al. 1972), C. septempunctata was virtually absent from experimental plots until 1980. Starting 1980, however, this species has become permanently established, and its relative abundance skyrocketed from 6.1% in 1980 to 100% in 1994 (Fig. 1). Concomitantly, the relative abundance of H. tredecimpunctata and C. transversoguttata decreased dramatically, with no C. transversoguttata encountered for periods up to several years. P. quatordecimpunctata was first observed in 1993, and H. axyridis was first observed in 1995. In the following years, both species became prominent members of the lady beetle community.
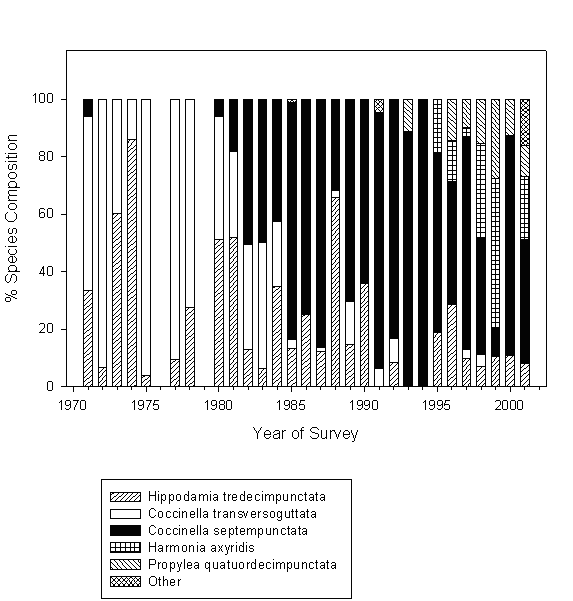
Fig. 1. Relative abundance of different lady beetle species in untreated potato plots in northern Maine in 1971-2001.
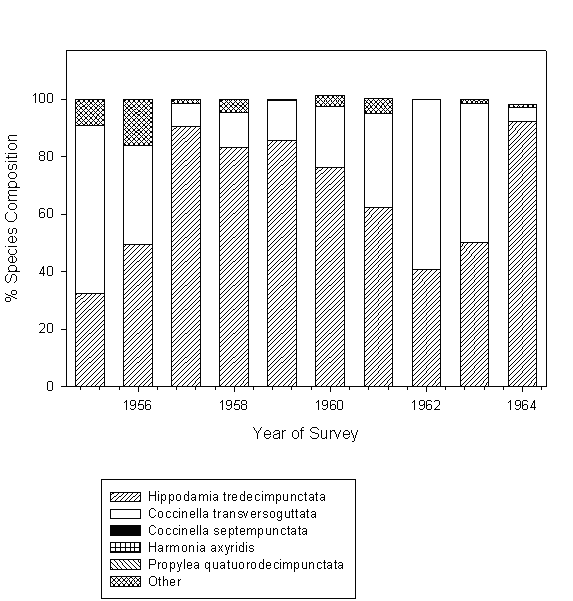
Fig. 2. Relative abundance of different lady beetle species in untreated potato plots in northern Maine in 1955-1964.
There were highly significant negative correlations between the abundance of native C. transversoguttata and all three species of exotic lady beetles (Table 1). Abundance of H. tredecimpunctata was negatively correlated with the abundance of C. septempunctata, but not the other two exotic species (possibly because H. tredecimpunctata populations had already been decimated at the time of their establishment in potato plots). In the same time, abundances of exotic species in most cases were positively correlated with each other (Table 1).
Table 1. Spearman’s rank correlation among densities of lady beetle populations in untreated potato plots in northern Maine.
|
|
C. transversoguttata |
C. septempunctata |
H. axyridis |
P. quatorodecimpunctata |
Other species |
| H. tredecimpunctata | Rs |
0.2574 |
-0.1241 |
-0.0840 |
-0.0149 |
-0.0760 |
|
P |
<.0001 |
0.0180 |
0.1100 |
0.7778 |
0.1483 |
|
|
|
|
|
|
|
C. transversoguttata |
Rs |
|
-0.2291 |
-0.1763 |
-0.1330 |
-0.0650 |
|
P |
|
<.0001 |
0.0007 |
0.0112 |
0.2165 |
|
|
|
|
|
|
|
C. septempunctata |
Rs |
|
|
0.0849 |
0.1740 |
0.0437 |
|
P |
|
|
0.1063 |
0.0009 |
0.4061 |
|
|
|
|
|
|
|
H. axyridis |
Rs |
|
|
|
0.4142 |
0.1425 |
|
P |
|
|
|
<.0001 |
0.0065 |
|
|
|
|
|
|
|
P. quatorodecimpunctata |
Rs |
|
|
|
|
0.0763 |
|
P |
|
|
|
|
0.1470 |
There were significant fluctuations in the diversity of lady beetle communities as evidenced both by the Berger-Parker species dominance index (F=5.13, df=28, 292, P<0.0001) and the Shannon diversity index (F=6.25, df=28, 292, P<0.0001). Species diversity, on average, increased following the establishment of the first exotic lady beetle, C. septempunctata, in 1980. Subsequent establishment of H. axyridis and P. quatordecimpunctata led to further elevation in species diversity (Table 2).
Table 2. Effect of C. septempunctata, H. axyridis, and P. quatordecimpunctata establishment on lady beetle community in untreated potato plots in northern Maine.
|
C. septempunctata |
|
H. axyridis and P. quatordecimpunctata |
||||||
|
Before |
After |
F3 |
P |
|
Before |
After |
F3 |
P |
Mean species diversity |
|
|
|
|
|
|
|
|
|
Berger index |
0.8733 |
0.7771 |
4.73 |
0.0305 |
|
0.7430 |
0.6743 |
14.27 |
0.0002 |
(SE) |
0.0152 |
0.0135 |
|
|
|
0.0221 |
0.0285 |
|
|
Shannon index |
0.2530 |
0.4372 |
8.02 |
0.0049 |
|
0.5004 |
0.6621 |
19.83 |
<0.0001 |
(SE) |
0.0268 |
0.0257 |
|
|
|
0.0415 |
0.0617 |
|
|
|
|
|
|
|
|
|
|
|
|
Mean lady beetle density1 |
|
|
|
|
|
|
|
|
|
All species |
0.0265 |
0.0212 |
8.26 |
0.0043 |
|
0.0230 |
0.0276 |
16.53 |
<0.0001 |
(SE) |
0.0026 |
0.0013 |
|
|
|
0.0024 |
0.0030 |
|
|
H. tredecimpunctata |
0.0120 |
0.0039 |
24.82 |
<0.0001 |
|
0.0057 |
0.0028 |
0.46 |
0.4995 |
(SE) |
0.0016 |
0.0004 |
|
|
|
0.0008 |
0.0006 |
|
|
| C. transversoguttata | 0.0144 |
0.0034 |
206.01 |
<0.0001 |
|
0.0083 |
0.0004 |
8.85 |
0.0031 |
(SE) |
0.0018 |
0.0005 |
|
|
|
0.0011 |
0.0002 |
|
|
|
|
|
|
|
|
|
|
|
|
Mean aphid density2 |
5.4828 |
2.7039 |
21.18 |
<0.0001 |
|
3.4154 |
0.6155 |
158.3 |
<0.0001 |
(SE) |
0.3177 |
0.1173 |
|
|
|
0.1722 |
0.0729 |
|
|
1 Per sampled plant (see text for sampling details).
2 Per sampled leaf (see text for sampling details).
3 Single degree of freedom orthogonal contrasts (Zar 1999).
Absolute abundance of lady beetles (all species pooled together) varied significantly among the years (F=10.84, df=28, 292, P<0.0001). Fewer beetles were encountered at the experimental plots after 1980 (Table 2). However, this difference was largely due to unusually high beetle populations in 1978 (Fig. 3). When that year was dropped from the analysis, the difference between those two time periods became statistically insignificant (F=0.49, P=0.4863). In the same time, more beetles were observed between 1980-95 than between 1995-2001 (Table 2). Establishment of C. septempunctata led to a pronounced reduction in density of H. tredecimpunctata and C. transversoguttata populations, while establishment of H. axyridis and P. quatordecimpunctata had a statistically significant impact only on C. transversoguttata (Table 2).
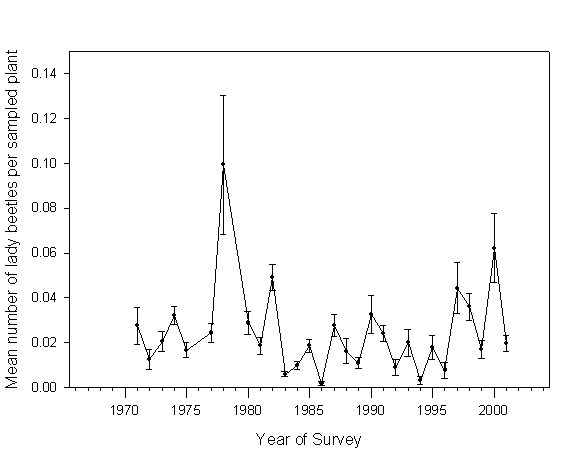
Fig. 3. Population density of lady beetles (all species combined) in untreated potato plots in northern Maine in 1971-2001.
Mean number of aphids infesting sampled leaves varied significantly among the years of the study (F=21.914, df=28, 292, P<0.0001). Noticeably fewer aphids were encountered at the experimental plots after establishment of exotic lady beetles, especially H. axyridis and P. quatordecimpunctata (Fig. 4; Table 2). Multiple regression analysis indicated that lady beetle density had a highly significant impact on aphid density (F=9.0, df=6, 356, P<0.0001), although the majority of observed variation was better explained by other factors (R2=0.13). Among all species, only densities of H. tredecimpunctata and H. axyridis were predictive of aphid density (Table 3). As evidenced by regression coefficients (Table 3), high densities of H. tredecimpunctata observed at experimental plots predicted that aphid densities would also be high, while high densities of H. axyridis predicted that aphid densities would be low.Table 3. Multiple regression results testing for effects of different lady beetle species on resident aphid populations in untreated potato plots in northern Maine.
Variable |
b |
SE |
t |
P |
Intercept |
176.86 |
7.58 |
23.34 |
<0.0001 |
| H. tredecimpunctata | 1783.69 |
451.33 |
3.95 |
<0.0001 |
C. transversoguttata |
410.64 |
386.94 |
1.06 |
0.2893 |
C. septempunctata |
-227.13 |
450.22 |
-0.50 |
0.6142 |
H. axyridis |
-6658.82 |
1791.26 |
-3.72 |
0.0002 |
P. quatorodecimpunctata |
-3710.91 |
2200.18 |
-1.69 |
0.0925 |
Other species |
-5041.58 |
4146.73 |
-1.22 |
0.2249 |
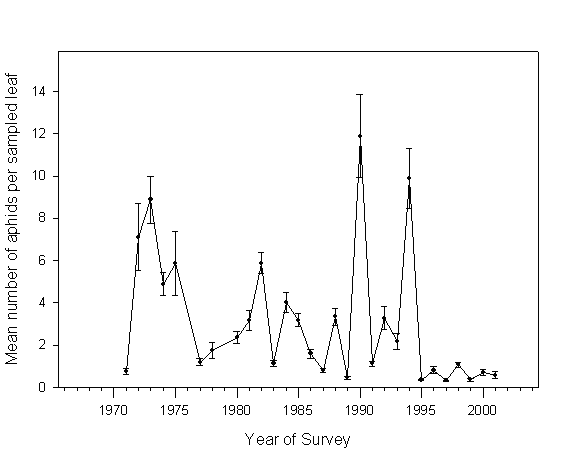
Fig. 4. Population density of potato-colonizing aphids in untreated potato plots in northern Maine in 1971-2001.
Discussion
Both density of lady beetle populations and relative abundance of different species comprising the lady beetle community varied significantly between the years of our study. Such a variation is quite common for this group of insects (Kieckhefer and Elliott 1990; Evans and Youssef 1992), and often makes it difficult to detect changes in their communities (Evans 2000). However, shifts in the species composition observed in our study appear to be consistent over long periods of time (Figs. 1 and 2). At first, the community previously consisting of native species became dominated by C. septempunctata. This is very similar to the situation observed by Wheeler and Hoebeke (1995) at a number of locations at northeastern North America, and by Elliott et al. (1996) in South Dakota. More recently, C. septempunctata domination was challenged by the other two exotic species, H. axyridis, and P. quatordecimpunctata. Brown and Miller (1998) reported comparable trend in West Virginia apple orchards. Therefore, it appears that changes observed in our study agree with broader changes occurring in lady beetle communities at a large geographic scale.
Interestingly, there was a highly significant positive correlation between the densities of H. axyridis, and P. quatordecimpunctata, as well as between the densities of P. quatordecimpunctata and C. septempunctata. This implies the possibility of biotic facilitation, a phenomenon when two or more invading species enhance each other’s survival and/or population size (Simberloff and Von Holle 1999). Biotic facilitation is probably more common in nature than previously thought (Simberloff and Von Holle 1999). However, our data are far from sufficient for making definite conclusions on this subject. It seems counter-intuitive that competitors would engage in facilitative interactions, and it is possible that the observed correlation is caused by some external factor having a similar impact on populations of different invading species.
Replacement of native lady beetles by C. septempunctata did not appear to cause significant reduction in the numbers of potato-colonizing aphids (Fig. 4), just as it was the case in the study by Evans (1991). In the same time, establishment of H. axyridis and P. quatordecimpunctata coincided with a dramatic decrease in the abundance of their prey (Fig. 4). Multiple regression analysis indicated that H. axyridis density had a significant influence on the density of aphid populations. No such effect was detected for P. quatordecimpunctata. Brown and Miller (1998) observed a similar suppression of pest aphids after H. axyridis had become a dominant species at their study sites, suggesting that H. axyridis is probably a superior biological control agent of aphid pests when compared to C. septempunctata.
Classical biological control via introduction of exotic natural enemies has come under increasing scrutiny because of its possible detrimental effects on non-target species (Howarth 1991; Simberloff and Stiling 1996; Lockwood 1993, 1996; Hopper 1998). Among other things, such negative interactions are caused by predation and competition for common resources, as was probably the case with the lady beetles observed in the present study. Although invasion of C. septempunctata, H. axyridis, and P. quatordecimpunctata is commonly thought to originate from inadvertent introductions rather than from deliberate releases (Day et al. 1994; Wheeler and Hoebeke 1995), our results still demonstrate the profound effects that exotic natural enemies may have on non-target native species. However, it is yet unknown if those effects have negative environmental consequences from the conservation standpoint.
Despite dramatic reduction in their relative abundances, both C. transversoguttata and H. tredecimpunctata persisted at low densities in potato plots after the establishment of exotic species (Fig. 1). As a result, the invasion increased the species diversity at the experimental plots (Table 3). Furthermore, potato is as exotic a species in northern Maine as C. septempunctata, H. axyridis, or P. quatordecimpunctata. In spite of usually being able to utilize a variety of different habitats, at least some aphidophagous lady beetle species have been shown to display preference towards certain environments (Honek 1985). Therefore, it has been argued that arrival of the invaders might simply cause ‘habitat compression’ for native lady beetles (Evans 2000). This entails retreat to ancestral habitats, where native species may have competitive advantage over the newcomers. Also, our study has been conducted using a small sampling unit at a single geographical location, and untreated plots were embedded within insecticide trial plots. All this does not allow us to make far-reaching conclusions solely based on our data. Further investigations on a larger spatial scale are required to fully assess the environmental impacts of exotic lady beetles. Knowing these impacts will improve our ability to evaluate the safety of entomophagous agents considered for introduction in biological control programs.
Acknowledgments
We thank Carrie Theriault for assistance with data entry. This is Publication No. 2628 of the Maine Agricultural and Forest Experiment Station.
References
Brown MW and Miller SS (1998) Coccinellidae (Coleoptera) in apple orchards of eastern West Virginia and the impact of invasion by Harmonia axyridis. Entomological News 109: 136-142
Chapin, JB and VA Brou (1991) Harmonia axyridis (Pallas), the third species of the genus to be found in the United States (Coleoptera: Coccinellidae). Proceedings of the Entomological Society of Washington 93: 630-635
Conover WJ and Iman RL (1981) Rank transformations as a bridge between parametric and nonparametric statistics. American Statistician 35: 124-129
Day, WH, Prokrym DR, Ellis DR, and Chianese RJ (1994) The known distribution of the predator Propylea quatordecimpunctata (Coleoptera: Coccinellidae) in the United States, and thoughts on the origin of this species and five other exotic lady beetles in eastern North America. Entomological News 105: 244-256
De Bach P and Rosen D (1991) Biological control by natural enemies. Cambridge University Press, Cambridge, 440 pp
Elliott N, Kieckhefer R, and Kauffman W (1996) Effects of an invading coccinellid on native coccinellids in an agricultural landscape. Oecologia 105: 537-544
Ellis DR, Prokrym DR, and Adams RG (1999) Exotic lady beetle survey in northeastern United States: Hippodamia variegata and Propylea quatordecimpunctata (Coleoptera: Coccinellidae). Entomological News 110: 73-84
Evans EW (1991) Intra versus interspecific interactions of ladybeetles (Coleoptera: Coccinellidae) attacking aphids. Oecologia 87: 401-408
Evans EW (2000) Morphology of invasion: body size patterns associated with establishment of Coccinella septempunctata (Coleoptera: Coccinellidae) in western North America. European Journal of Entomology 97: 469-474
Evans EW and Youseff NN (1992) Numerical responses of aphid predators to varying prey density among Utah alfalfa fields. Journal of Kansas Entomological Society 65: 30-38
Gordon RD (1985) The Coccinellidae (Coleoptera) of America north of Mexico. Journal of the New York Entomological Society 93: 1-912
Hironori, Y and Katsuhiro S (1997) Cannibalism and interspecific predation in two predatory ladybirds in relation to prey abundance in the field. Entomophaga 42: 153-163
Hoebeke ER and Wheeler AG (1996) Adventive lady beetles (Coleoptera: Coccinellidae) in the Canadian Maritime provinces, with new eastern U.S. records of Harmonia quadripunctata. Entomological News 281-290
Honek A (1985) Habitat preferences of aphidophagous coccinellids (Coleoptera). Entomophaga 30: 253-264
Hopper KR (1998) Assessing and improving the safety of introductions for biological control. Phytoprotection 79(S): 84-93
Howarth F G (1991) Environmental impacts of classical biological control. Annual Review of Entomology 36: 485-509
Kajita Y, Takano F, Yasuda H, and Agarwala BK (2000) Effects of indigenous ladybird species (Coleoptera: Coccinellidae) on the survival of an exotic species in relation to prey abundance. Applied Entomology and Zoology 35: 473-479
Kiekhefer RW and Elliott NC (1990) A 13-year survey of the aphidophagous Coccinellidae in maize fields in eastern South Dakota. Canadian Entomologist 122: 579-581
Lockwood JA (1993) Environmental issues involved in biological control of rangeland grashoppers (Othoptera: Acrididae) with exotic agents. Environmental Entomology 22: 503-518
Lockwood JA (1996) The ethics of biological control: understanding the moral implications of our most powerful ecological technology. Agriculture and Human Values 13: 2-19
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, and Bazzaz F.A (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecological Applications 10: 689-710
Magurran AE (1988) Ecological diversity and its measurement. Princeton University Press, Princeton, New Jersey, 179 pp
Obrycki JJ, Giles KL, and Ormond AM (1998) Experimental assessment of interactions between larval Coleomegilla maculata and Coccinella septempunctata (Coleoptera: Coccinellidae) in field cages. Environmental Entomology 27: 1280-1288
SAS Institute, Inc. (1999) SAS OnLine Doc, Version 8. Cary, North Carolina
Schaefer, PW, Dysart RJ, and Specht HB (1987) North American distribution of Coccinella septempunctata (Coleoptera: Coccinellidae) and its mass appearance in coastal Delaware. Environmental Entomology 16: 368-373
Shands, WA, Simpson GW, Wave HE, and Gordon CC (1972) Importance of arthropod predators in controlling aphids on potatoes in northeastern Maine. University of Maine (Orono) Technical Bulletin 54
Simberloff D. and Stiling P (1996) How risky is biological control? Ecology 77:1965-1974
Simberloff D, Von Holle B (1999) Positive interactions of nonindigenous species: invasional meltdown? Biological Invasions 1, 21-32
Wheeler AG and Hoebeke ER (1995) Coccinella novemnotata in northeastern North America: historical occurrence and current status (Coleoptera: Coccinellidae). Proceedings of the Entomological Society of Washington 97: 701-716
Wright RJ and DeVries TA (2000) Species composition and relative abundance of Coccinellidae (Coleoptera) in south central Nebraska field crops. Journal of Kansas Entomological Society 73: 103-111
Zar JH (1999) Biostatistical Analysis, 4th Edition. Prentice Hall, Upper Saddle River, New Jersey, 663 pp