Colorado Potato Beetle Response to Soil Amendments: A Case in Support of the Mineral Balance Hypothesis?
Andrei Alyokhin, Gregory Porter*, Eleanor Groden, and Francis Drummond
Department of Biological Sciences, University of Maine, 5722 Deering Hall, Orono, ME 04469
* Department of Plant, Soil, and Environmental Sciences, University of Maine, 5722 Deering Hall, Orono, ME 04469
This peer-refereed invited review article has been first published by Agriculture, Ecosystems, and Environment (Volume 109, Pages 234-244). The original publication is available at doi:10.1016/j.agee.2005.03.005. Reproduced with permission of the publisher. Please send me an e-mail if you want a PDF reprint of this article in its original journal format.
ABSTRACT
The mineral balance hypothesis (Phelan et al., 1996) suggests that the organic matter and microbial activity associated with organically managed soils afford a buffering capability to maintain nutrient balance in plants. An optimal nutrient balance, in turn, results in both good plant growth and resistance to herbivory. Effects of soil amendment practices on Colorado potato beetle populations in potato fields and their interactions with crop rotation and two pest management approaches were investigated in the present study. Beetle densities were generally lower in plots receiving manure soil amendments in combination with reduced amounts of synthetic fertilizers compared to plots receiving full rates of synthetic fertilizers, but no manure. Crop rotation and pest management approaches had little or no effect. Unlike beetle abundance, plant height and canopy cover were comparable between plots receiving manure and synthetic fertilizer. Furthermore, tuber yields were higher in manure-amended plots. In direct accordance with the mineral balance hypothesis, there was a dramatic dissimilarity in mineral composition of potato leaves collected from manure-amended and synthetic fertilizer-treated plots. Overall, there were differences in concentrations of nitrogen, calcium, magnesium, phosphorus, aluminum, boron, copper, iron, manganese, and zinc. Boron concentration was most dramatically affected by the soil amendment. Mineral content of potato leaves explained 40-57% of the variation in the Colorado potato beetle populations observed among the experimental plots.
Key words: mineral balance, plant nutrients, soil fertilization, manure, insect herbivory, Colorado potato beetle, sustainable agriculture
Introduction
One of the foundations of organic farming is an assumption that the organic production systems create a generally unfavorable environment for pest populations (Oelhaf, 1978; Beanland et al., 2003). Indeed, insect populations are often comparable between organic and conventional fields, even though organically certified insecticides are usually less effective than their conventional counterparts (Feber et al., 1997; Gallandt et al., 1998; Letourneau and Goldstein, 2001; Delate et al., 2003). Furthermore, plants grown on organically managed soils fertilized with manure and compost have been shown to be less favorable hosts for phytophagous insects than plants grown on conventionally managed soils fertilized with synthetic fertilizers (Eigenbrode and Pimentel, 1988; Phelan et al., 1995, 1996).
In an attempt to explain apparent reductions in pest pressure in organic systems, Phelan et al. (1996) and Phelan (1997) formulated the mineral balance hypothesis. This hypothesis suggests that the organic matter and microbial activity associated with organically managed soils afford a buffering capability to maintain nutrient balance in plants. An optimal nutrient balance results in both good plant growth and resistance to herbivory. In contrast, crops growing in soils without these biologically based buffering capabilities are more likely to take up either excess or insufficient levels of certain nutrients. In some instances, resulting imbalance in the ratio of certain mineral nutrients may result in rapid plant growth. However, affected plants may have their primary and/or secondary metabolism impaired, thus compromising their ability to resist or tolerate insect damage. A number of experimental studies (Clancy et al., 1988; Clancy, 1992; Phelan et al., 1996; Busch and Phelan, 1999; Beanland et al., 2003) provided empirical support to the formulated hypothesis.
Colorado potato beetle, Leptinotarsa decemlineata (Say), is the most important insect defoliator of potatoes (Weber and Ferro, 1994). None of the control techniques developed against this pest during the past 135 years has provided long-term protection of potato crops (Casagrande, 1987), and the beetle continues to be a major threat to potato production. High fecundity, a diverse and flexible life history, and a remarkable ability to develop insecticide resistance make Colorado potato beetle management a challenging task (Weber and Ferro, 1994). If soil management practices decrease beetle pressure, they may provide a valuable addition to the chemically skewed arsenal that is currently used to control this pest.
Materials and Methods
The study was conducted during the 1999 – 2003 growing seasons at the Maine Agricultural and Forest Research Station’s Aroostook Research Farm in Presque Isle, Maine. The land used for the study had a long history of commercial and research potato production. The soil for the entire site was a gravely, well-drained Caribou loam (fine-loamy, mixed, frigid, Typic Haplorthods) that contained extensive coarse fragments (Porter, 1996; Gallandt et al., 1998).
The site was separated into 96 plots arranged in four statistical blocks. Thirty-two plots were in potato production during each year of the study. The remaining plots were planted to rotation crops as described below. Each plot was 41.0 m long and 14.6 m wide. Potatoes in all plots were planted in late May using a “pick”-type planter at the depth of 5-10 cm. Seed tubers were hand cut into ca. 50 g pieces prior to planting. The distance between the rows was 90 cm, and tuber spacing within the rows was 23 cm. Blended fertilizer was applied in a band 5 cm below and 5 cm to the side of the seed pieces at planting. Potatoes were ridged »4 weeks after planting.
The study consisted of two experiments, both of which were laid out following a randomized complete block, fully factorial design. Treatments tested in each of the experiments are described in the following sections. The main objective of Experiment 1 was to determine effects of different rotation schedules and their interaction with soil amendments on pest populations and crop development. Crop rotation and soil amendment system were the main factors randomly assigned within each of the four blocks. Pests on all plots in this experiment were managed using a conventional IPM approach.
The main objective of Experiment 2 was to determine effects of different pest management systems and their interaction with soil amendments on pest populations and crop development. Correspondingly, pest management system (conventional IPM or biorational IPM) and the soil amendment system were the main factors. All plots in this experiment were on the integrated rotation schedule. Because of the space constraints, plots designated as conventional IPM plots in the second experiment were the same plots designated as plots on the integrated rotation schedule in the first experiment. There were no rotation schedule comparisons within the biorational IPM system. Therefore, the two experiments were not truly independent from each other.
Half of the plots set up in the present study received only synthetic fertilizer, while the other half was amended with raw cow manure mixed with sawdust as bedding material. Nutrient needs of the synthetic system were met with chemical fertilizer applied at recommended rates (1344 kg/ha of 10-10-10 applied at planting and 56 kg/ ha of nitrogen side dressed at tuber initiation). Additional potash fertilizer was also broadcast in the spring based on soil test recommendations. Nutrients in the manure-amended system were provided by raw cow manure (rates adjusted to provide a current-season average of 107 kg/ha of plant available N based on the analysis conducted by the University of Maine Analytical Laboratory), residual soil fertility, and an at-planting application of ammonium sulfate fertilizer (78 kg/ha of actual N) to make up the balance of N fertilizer needs. The manure-amended system had been receiving annual applications of waste potato compost (22000 kg/ha, providing ca. 13 kg/ha of plant available nitrogen) and cattle manure (44000 kg/ha, providing ca. 72 kg/ha of plant available nitrogen) prior to primary tillage each spring from 1991-98. The rotation crop used from 1991-98 in the manure-amended system during that period was a green manure crop consisting of peas, oats, and hairy vetch. As a result, fertility, organic matter levels, and physical properties of the soils in synthetically fertilized and manure-amended plots were already different at the beginning of this study (Porter and McBurnie, 1996; Gallandt et al., 1998). During the present study, soil organic matter was about 1.7 times higher on the amended plots than on the synthetically fertilized plots (4.7% vs. 2.8% in Experiment 1, and 4.5% vs. 2.7% in Experiment 2).
Three crop rotations were compared. Those were: (a) two-year standard (potatoes : barley), (b) four-year intensive (potatoes : soybean : potatoes : barley) and (c) four-year integrated (potatoes : soybean : barley : mixed alfalfa and timothy stand).
Two different integrated pest management approaches to controlling Colorado potato beetles were evaluated. Both were based on the recommendations developed by the University of Maine Cooperative Extension (Dwyer et al., 1997), although the thresholds have been modified to provide adequate control in this particular experimental setting (Drummond and Groden, 1996; Gallandt et al., 1998). For the Colorado potato beetles, the threshold densities were 1 adult, 8 small larvae, or 3 large larvae per plant in the conventional IPM system, and 0.5 adults, 1.5 small larvae, or 1.5 large larvae per plant for the biorational IPM system.
The conventional IPM system relied on foliar applications of imidacloprid (52.6 g ai/ha, Provado®) for insect control. The biorational IPM system utilized foliar-applied Bacillus thuringiensis (79 g ai/ha, Foil®) and the fungal pathogen, Beauveria bassiana (5 x 1013 B. bassiana conidia/ha, Mycotrol®) for control of larvae. Rotenone plus PBO (280 g ai/ha, Rotacide®) was applied for control of summer adults when needed. Methamidophos (1680 g ai/ha, Monitor®) was used to control aphids in the conventional IPM system, and pyrethrin (52.6 g ai/ha, Pyrenone®) and pymetrozine (96 g ai/ha, Fulfil®) were used in the biorational IPM system. Fungal diseases in the conventional IPM plots were controlled with foliar applications of chlorothalonil (1260 g ai/ha, Bravo®) and EBDCs (1680 g ai/ha, Mancozeb®, Penncozeb®, or Maneb®). In the biorational IPM system, fungal diseases were controlled by foliar applications of chlorothalonil (1260 g ai/ha, Bravo®) and strobilurin (104 g ai/ha, Quadris®).
Thirty potato plants were randomly selected within each plot for weekly visual examinations. The number of Colorado potato beetles was recorded on a whole-plant basis, with adults, egg masses, and small (first and second instars combined) and large (third and fourth instars combined) larvae recorded separately.
All plant sampling was done only during the four growing seasons in 2000-03. No plant data were taken in 1999. Plant height was measured four times at weekly intervals starting the second week in July. Ten plants were randomly selected at each plot and measured with a meter stick from the top of the row to the top of the plant. Additionally, percent ground cover was visually estimated during the second week of July in each plot. To minimize human error, three readings per plot were taken, and the average was used in the analysis.
Thirty most recently expanded leaves (typically 4th to 5th from top of plant) were collected at random from each plot during the last week of July or the first week of August. Collected leaves were washed in deionized water, dried at 60°C, ground, and then submitted to the University of Maine Analytical Laboratory for nutrient analysis.
Potato vines were killed during the second week of September using a flail-type mower in the biorational IPM plots, and using commercial desiccant (Reglone®) in the conventional IPM plots. Yields were then determined from the four center rows of each plot (25% of the plot area). The tubers were excavated with a two-row potato digger, collected by hand, and weighed.
To reduce the number of zeroes, insect counts were pooled together for all 30 plants sampled within the same plot. Data normality was tested before the analyses at 0.01 level of significance using Kolmogorov-Smirnov tests (PROC UNIVARIATE, SAS Institute, 1999). When necessary, the data were transformed using rank transformations (Conover and Iman, 1981). Means and standard errors were calculated from the untransformed data. Repeated measures analyses of variance (PROC GLM, SAS Institute, 1999) were used to compare insect populations and plant heights throughout the season during each year of the study. To compensate for heterogeneity in both variances and covariances, degrees of freedom of F-tests for within-subject effects were adjusted using the Huynh-Feldt epsilon (Huynh and Feldt, 1970). Tuber yields, percent cover, and nutrient content of the sampled leaves were compared using factorial analyses of variance (PROC GLM, SAS Institute, 1999). Multiple regression analyses (PROC REG, SAS Institute, 1999) were used to analyze the effects of leaf nutrients on the Colorado potato beetle population densities. Data from all 96 plots (both experiments combined) were pooled together for 2000-03 and analyzed as a single data set.
Results
Soil amendment appeared to be the major factor affecting Colorado potato beetle density in experimental plots. Beetle densities in both experiments were almost always lower in the plots receiving manure (Fig. 1 and Fig. 2). Unlike soil amendment, rotation sequence had relatively little effect on beetle populations. In 1999, significantly (F2,5=6.33; P=0.0427) fewer overwintered adults were detected in plots on the two-year rotation (0.008±0.003 [mean±SE] insects/plant) compared to plots on the integrated (0.024±0.006 insects/plant) or intensive (0.029±0.01 insects/plant) rotation schedules. However, that was the first year of the experiment, and the observed differences could hardly be attributed to the rotation itself. In 2003, significantly (F2,4=29.48; P=0.0056) more large larvae were encountered in plots on the two-year rotation (1.72±0.35 insects/plant) than in plots on the integrated (1.51±0.29 insects/plant) or intensive (1.45±0.21 insects/plant) rotation schedules. In all other instances, beetle densities were similar among the rotations. None of the interactions between the soil amendment and crop rotation were statistically significant.
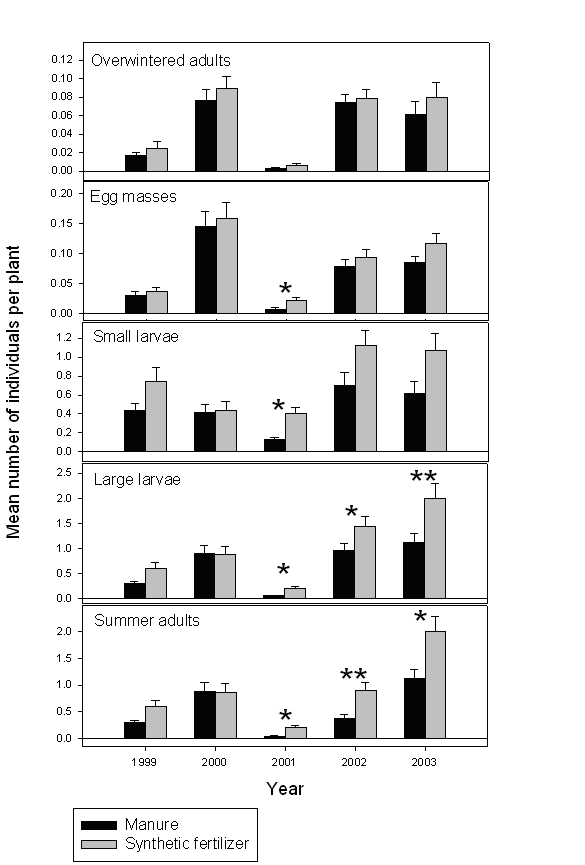
Figure 1. Densities of the Colorado potato beetle life stages in manure-amended and synthetic fertilizer-treated plots on different rotation schedules (Experiment 1). Significant differences between the treatments are indicated with * (P<0.05) and ** (P<0.01).
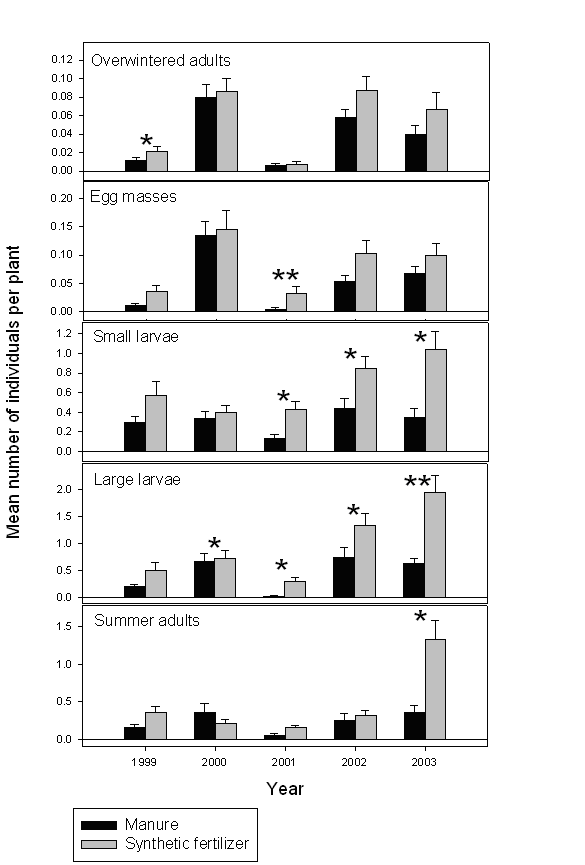
Figure 2. Densities of the Colorado potato beetle life stages in manure-amended and synthetic fertilizer-treated plots under conventional and biorational integrated pest management systems (Experiment 2). Significant differences between the treatments are indicated with * (P<0.05) and ** (P<0.01).
In 2000, biorational plots contained significantly (F1,3=28.18; P=0.0130) fewer summer adults (0.05±0.01 beetles/plant) than conventional plots (0.51±0.12 insects /plant). Similarly, in 2002 significantly lower densities of overwintered adults (0.041±0.010 insects/plant vs. 0.064±0.018 insects/plant; F1,3=21.89; P=0.0184), large larvae (0.85±0.18 insects/plant vs. 1.24±0.23 insects/plant; P=0.0075), and summer adults (0.05±0.02 insects/plant vs. 0.51±0.10 insects/plant; F1,3=46.06; P=0.0065) were recorded in the biorational plots. Also, there was a significant interaction (F1,3=28.18; P=0.0130) between the effects of pest management system and soil amendment on the density of large larvae. Manure-amended biorational plots had the fewest larvae (0.30±0.13 insects/plant), followed by manure-amended conventional plots (1.21±0.32 insects/plant), synthetic fertilizer conventional plots (1.28±0.34 insects/plant), and synthetic fertilizer biorational plots (1.40±0.31 insects/plant).
Both soil amendment and crop rotation affected the height of potato plants, but only during two years of the study (Table 1). Also, in 2003 plants were slightly taller (F3,6=6.25; P=0.0465) in biorational plots (52.5±2.2 cm) than in the conventional IPM plots (50.4±2.0 cm).
In Experiment 1, canopy cover at the time of measurement was significantly greater (F1,6=11.38; P=0.015) in plots treated with synthetic fertilizer (40.4±1.2%) compared to the plots amended with manure (36.2±1.0%) in 2002. Also, that year canopy cover was higher (F2,6=9.59; P=0.0135) in plots on the 2-year rotation schedule (41.6±1.4%) compared to plots on 4-year integrated (35.4±1.1%) or intensive (37.9±1.3%) rotation schedules. In Experiment 2, canopy cover was marginally higher (F1,4=10.26; P=0.0492) in manure-amended plots (52.2±1.2%) than in synthetic fertilizer plots (47.7±1.2%) in 2000. Significantly more tubers were harvested from manure-amended plots in Experiment 1 during four out of five years of the study (Fig. 3). Rotation effects were not significant, and neither were the interactions between soil amendment and rotation.
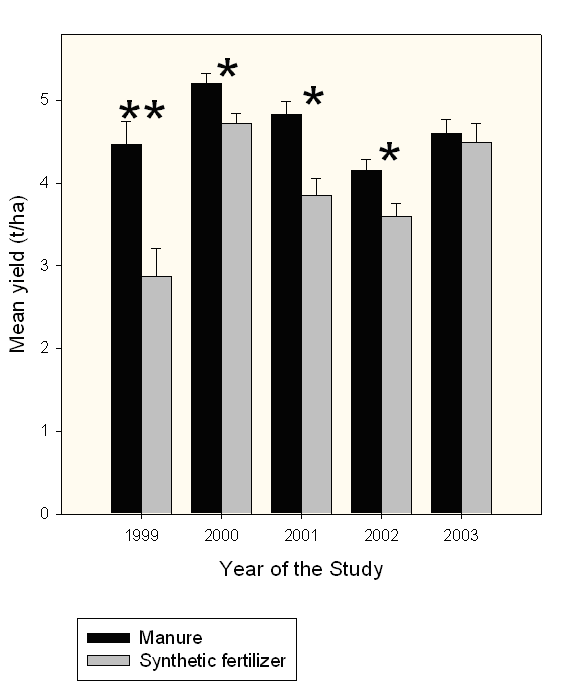
Figure 3. Yields of potato tubers at manure- and synthetic fertilizer-amended plots on different rotation schedules (Experiment 1). Significant differences between the treatments are indicated with * (P<0.05) and ** (P<0.01).
Soil amendment significantly affected both macro- and micronutrient concentration in potato foliage. Effect of crop rotation was less pronounced, but we still detected occasional significant differences (Tables 2-5). Multiple regression analyses indicated that densities of all Colorado potato beetle life stages were significantly affected by the leaf mineral concentration, and this accounted for a considerable proportion of the observed variation in Colorado potato beetle density (Table 6). Boron and phosphorus appeared to have consistently negative effect on beetle numbers, while zinc had a consistently positive effect.
Table 1. Heights of potato plants as affected by soil amendment and rotation schedule (mean±SE). Statistically significant effects are indicated as * (P<0.05) and ** (P<0.01). Factors that were tested in the study but are not listed in this table were not statistically significant (P>0.05).
|
|
2000 |
2001 |
2002 |
2003 |
Experiment 1 |
|
|
|
|
|
|
Manure |
44.19±1.89 |
56.37±1.64** |
43.39±2.05* |
51.59±1.66 |
|
Synthetic fertilizer |
43.30±2.06 |
51.92±1.42 |
45.05±1.95 |
50.11±1.64 |
|
|
|
|
|
|
|
Integrated 4-year rotation |
44.85±2.59 |
54.09±1.92** |
42.71±2.49* |
50.36±1.99 |
|
Intensive 4-year rotation |
43.20±2.28 |
53.39±1.88 |
44.64±2.42 |
50.73±2.01 |
|
Two-year rotation |
43.19±2.42 |
54.96±1.95 |
45.30±2.48 |
51.46±2.09 |
|
|
|
|
|
|
|
|
|
|
|
|
Experiment 2 |
|
|
|
|
|
|
Manure |
45.76±2.70 |
56.35±1.99** |
41.74±2.45 |
52.08±2.15 |
|
Synthetic fertilizer | 44.17±2.45 |
51.75±1.87 |
43.78±2.55 |
50.73±2.01 |
Table 2. Concentration of macronutrients (mean±SE, % dry weight) in potato leaves collected from manure-amended and synthetic fertilizer-treated plots on different rotation schedules (Experiment 1). Statistically significant effect of soil amendment are indicated as A* (P<0.05) and A** (P<0.01). Statistically significant effects of crop rotation schedule are indicated as R* (P<0.05) and R** (P<0.01).
Year |
Mineral |
Effect | Integrated Rotation |
|
Intensive Rotation |
|
Two-year Rotation |
|||
|
|
|
Manure |
Synthetic |
|
Manure |
Synthetic |
|
Manure |
Synthetic |
2000 |
N |
A* |
5.18±0.24 |
5.81±0.16 |
|
5.43±0.08 |
5.83±0.22 |
|
5.52±0.15 |
5.74±0.12 |
|
Ca |
ns |
0.90±0.08 |
0.87±0.06 |
|
0.93±0.01 |
0.81±0.03 |
|
0.89±0.05 |
0.87±0.03 |
|
K |
ns |
2.98±0.17 |
3.27±0.18 |
|
3.12±0.17 |
3.42±0.28 |
|
3.48±0.17 |
3.39±0.02 |
|
Mg |
ns |
0.58±0.04 |
0.63±0.03 |
|
0.62±0.02 |
0.64±0.05 |
|
0.59±0.01 |
0.62±0.03 |
|
P |
A* |
0.36±0.01 |
0.40±0.02 |
|
0.35±0.01 |
0.40±0.01 |
|
0.38±0.01 |
0.40±0.01 |
|
|
|
|
|
|
|
|
|
|
|
2001 |
N |
ns |
6.20±0.12 |
6.37±0.09 |
|
5.91±0.11 |
6.11±0.19 |
|
6.15±0.13 |
6.16±0.18 |
|
Ca |
ns |
0.90±0.12 |
0.88±0.08 |
|
0.94±0.07 |
0.87±0.11 |
|
0.85±0.09 |
0.96±0.08 |
|
K |
ns |
4.06±0.39 |
3.99±0.24 |
|
4.11±0.34 |
4.12±0.40 |
|
4.09±0.22 |
4.28±0.17 |
|
Mg |
ns |
0.61±0.04 |
0.62±0.01 |
|
0.61±0.02 |
0.58±0.05 |
|
0.60±0.04 |
0.68±0.02 |
|
P |
A* |
0.49±0.02 |
0.46±0.02 |
|
0.51±0.04 |
0.44±0.01 |
|
0.52±0.03 |
0.44±0.02 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2002 |
N |
ns |
6.38±0.07 |
6.43±0.08 |
|
6.01±0.15 |
5.88±0.26 |
|
6.19±0.09 |
5.86±0.32 |
|
Ca |
ns |
1.19±0.06 |
1.24±0.11 |
|
1.27±0.06 |
1.53±0.10 |
|
1.26±0.08 |
1.31±0.09 |
|
K |
ns |
5.27±0.33 |
4.77±0.12 |
|
5.59±0.18 |
5.02±0.40 |
|
5.32±0.31 |
5.21±0.24 |
|
Mg |
A**, R* |
0.76±0.03 |
0.85±0.04 |
|
0.79±0.04 |
1.00±0.06 |
|
0.82±0.04 |
0.89±0.13 |
|
P |
ns |
0.35±0.02 |
0.41±0.02 |
|
0.38±0.04 |
0.37±0.03 |
|
0.38±0.04 |
0.35±0.01 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2003 |
N |
A**, R** |
5.08±0.09 |
5.34±0.07 |
|
4.74±0.04 |
5.11±0.05 |
|
5.07±0.05 |
5.06±0.05 |
|
Ca |
ns |
0.64±0.04 |
0.70±0.03 |
|
0.67±0.02 |
0.70±0.03 |
|
0.59±0.04 |
0.66±0.01 |
|
K |
R* |
3.67±0.10 |
3.91±0.14 |
|
3.88±0.07 |
4.40±0.17 |
|
4.30±0.08 |
4.27±0.12 |
|
Mg |
ns |
0.49±0.04 |
0.49±0.02 |
|
0.46±0.03 |
0.47±0.02 |
|
0.43±0.01 |
0.47±0.01 |
|
P |
A** |
0.32±0.01 |
0.39±0.01 |
|
0.32±0.00 |
0.39±0.01 |
|
0.36±0.01 |
0.39±0.01 |
Table 3. Concentration of micronutrients (mean±SE, parts per million dry weight) in potato leaves collected from manure-amended and synthetic fertilizer-treated plots on different rotation schedules (Experiment 1). Statistically significant effect of soil amendment are indicated as A* (P<0.05) and A** (P<0.01). Statistically significant effects of crop rotation schedule are indicated as R* (P<0.05) and R** (P<0.01).
Year |
Mineral |
Effect | Integrated Rotation |
|
Intensive Rotation |
|
Two-year Rotation |
|||
|
|
|
Manure |
Synthetic |
|
Manure |
Synthetic |
|
Manure |
Synthetic |
2000 |
Al |
ns |
33.75±7.04 |
37.35±4.66 |
|
54.28±4.84 |
35.88±5.04 |
|
40.73±8.65 |
47.00±6.95 |
|
B |
A** |
20.43±0.97 |
10.44±1.36 |
|
20.78±1.21 |
10.74±0.59 |
|
23.10±0.36 |
10.25±0.94 |
|
Cu |
A* |
12.20±0.77 |
13.85±0.55 |
|
11.65±0.55 |
14.08±0.65 |
|
11.70±0.85 |
14.15±0.57 |
|
Fe |
ns |
105.78±5.61 |
106.28±3.88 |
|
108.45±9.18 |
103.45±6.04 |
|
114.20±10.41 |
107.75±2.39 |
|
Mn |
ns |
149.53±21.20 |
195.25±17.05 |
|
170.00±10.61 |
221.50±32.85 |
|
195.25±51.92 |
209.00±32.99 |
|
Zn |
ns |
20.60±0.79 |
22.20±2.09 |
|
20.55±1.55 |
25.85±0.60 |
|
28.03±4.53 |
22.20±1.60 |
|
|
|
|
|
|
|
|
|
|
|
2001 |
Al |
ns |
22.53±7.03 |
21.33±3.65 |
|
20.60±4.64 |
29.10±11.53 |
|
13.97±4.54 |
25.63±3.95 |
|
B |
A** |
25.63±1.13 |
17.18±1.12 |
|
27.70±0.44 |
17.15±0.61 |
|
25.60±0.98 |
18.05±0.86 |
|
Cu |
ns |
14.13±0.51 |
13.23±0.37 |
|
14.93±0.90 |
13.80±0.63 |
|
14.85±0.78 |
13.85±0.78 |
|
Fe |
ns |
108.25±1.80 |
116.50±5.55 |
|
110.25±4.31 |
118.30±11.88 |
|
107.08±6.76 |
106.48±5.19 |
|
Mn |
A** |
74.15±6.09 |
106.75±2.81 |
|
78.53±3.91 |
152.25±23.39 |
|
86.93±8.67 |
154.50±22.07 |
|
Zn |
ns |
21.85±1.79 |
21.50±1.10 |
|
22.73±1.50 |
22.83±0.58 |
|
21.83±1.73 |
21.55±0.90 |
|
|
|
|
|
|
|
|
|
|
|
2002 |
Al |
ns |
57.78±9.30 |
61.38±8.88 |
|
52.70±4.92 |
62.83±3.43 |
|
52.93±4.06 |
59.30±3.27 |
|
B |
A** |
28.38±0.93 |
16.28±2.13 |
|
32.33±0.63 |
14.28±1.60 |
|
32.08±1.44 |
17.18±3.04 |
|
Cu |
A**, R** |
15.65±1.38 |
20.25±1.25 |
|
14.00±1.45 |
15.68±0.48 |
|
13.50±1.70 |
14.68±0.49 |
|
Fe |
ns |
104.55±7.59 |
118.33±11.40 |
|
107.50±4.86 |
118.75±4.87 |
|
111.48±6.38 |
116.25±4.70 |
|
Mn |
A** |
111.55±10.36 |
146.25±13.42 |
|
145.50±12.55 |
219.25±14.30 |
|
102.33±5.30 |
179.48±44.87 |
|
Zn |
ns |
33.05±5.56 |
31.03±2.40 |
|
29.65±3.15 |
38.58±11.01 |
|
46.45±18.86 |
27.38±4.71 |
|
|
|
|
|
|
|
|
|
|
|
2003 |
Al |
ns |
43.30±2.39 |
35.33±1.93 |
|
56.93±20.88 |
36.68±4.32 |
|
34.65±3.78 |
34.70±3.13 |
|
B |
A** |
19.33±0.39 |
12.75±0.57 |
|
20.55±0.22 |
13.45±0.73 |
|
21.60±0.70 |
13.68±0.45 |
|
Cu |
A** |
11.35±0.82 |
12.78±0.92 |
|
11.21±1.16 |
13.03±0.78 |
|
10.26±0.41 |
13.90±0.32 |
|
Fe |
A* |
81.85±1.49 |
88.48±2.67 |
|
78.78±1.51 |
82.75±1.20 |
|
79.25±1.17 |
82.63±2.08 |
|
Mn |
A**, R** |
128.50±9.08 |
182.25±16.73 |
|
125.25±10.24 |
261.00±17.83 |
|
140.00±5.60 |
262.75±18.45 |
|
Zn |
A* |
21.38±1.19 |
22.43±2.26 |
|
20.28±0.78 |
25.15±1.20 |
|
21.98±0.71 |
24.98±2.07 |
Table 4. Concentration of macronutrients (mean±SE, % dry weight) in potato leaves collected from manure-amended and synthetic fertilizer-treated plots under conventional and biorational integrated pest management systems (Experiment 2). Statistically significant effect of soil amendment are indicated as A* (P<0.05) and A** (P<0.01). Statistically significant effects of pest management system are indicated as PM* (P<0.05) and PM** (P<0.01). Data for 2001 and 2002 are omitted because none of the differences between the treatments were statistically significant.
Year |
Mineral |
Effect | Biorational IPM |
|
Conventional IPM |
||
|
|
|
Manure |
Synthetic |
|
Manure |
Synthetic |
2000 |
N |
A** |
5.65±0.19 |
6.15±0.24 |
|
5.18±0.24 |
5.81±0.16 |
|
Ca |
ns |
0.92±0.07 |
1.03±0.09 |
|
0.90±0.08 |
0.87±0.06 |
|
K |
PM* |
3.79±0.12 |
3.56±0.19 |
|
2.98±0.17 |
3.27±0.18 |
|
Mg |
ns |
0.61±0.05 |
0.75±0.04 |
|
0.58±0.04 |
0.63±0.03 |
|
P |
ns |
0.40±0.01 |
0.40±0.02 |
|
0.36±0.01 |
0.40±0.02 |
|
|
|
|
|
|
|
|
2003 |
N |
A* |
5.20±0.11 |
5.27±0.07 |
|
5.08±0.09 |
5.34±0.07 |
|
Ca |
A*, PM* |
0.59±0.04 |
0.64±0.02 |
|
0.64±0.04 |
0.70±0.03 |
|
K |
ns |
4.17±0.17 |
4.22±0.29 |
|
3.67±0.10 |
3.91±0.14 |
|
Mg |
ns |
0.43±0.03 |
0.48±0.04 |
|
0.49±0.04 |
0.49±0.02 |
|
P |
A** |
0.35±0.01 |
0.39±0.01 |
|
0.32±0.01 |
0.39±0.01 |
Table 5. Concentration of micronutrients (mean±SE, parts per million dry weight) in potato leaves collected from manure-amended and synthetic fertilizer-treated plots under conventional and biorational integrated pest management systems (Experiment 2). Statistically significant effect of soil amendment are indicated as A* (P<0.05) and A** (P<0.01). Statistically significant effects of pest management system are indicated as PM* (P<0.05) and PM** (P<0.01).
Year |
Mineral |
Effects | Biorational IPM |
|
Conventional IPM |
||
|
|
|
Manure |
Synthetic |
|
Manure |
Synthetic |
2000 |
Al |
ns |
72.83±29.04 |
41.40±5.67 |
|
33.75±7.04 |
37.35±4.66 |
|
B |
A**, PM* |
24.38±0.60 |
10.78±0.29 |
|
20.43±0.97 |
10.44±1.36 |
|
Cu |
ns |
15.73±1.16 |
16.58±1.42 |
|
12.20±0.77 |
13.85±0.55 |
|
Fe |
ns |
157.25±38.88 |
120.75±6.12 |
|
105.78±5.61 |
106.28±3.88 |
|
Mn |
ns |
104.80±15.21 |
134.00±15.83 |
|
149.53±21.20 |
195.25±17.05 |
|
Zn |
ns |
19.93±1.01 |
18.10±1.18 |
|
20.60±0.79 |
22.20±2.09 |
|
|
|
|
|
|
|
|
2001 |
Al |
ns |
49.82±25.96 |
48.05±10.87 |
|
22.53±7.03 |
21.33±3.65 |
|
B |
A** |
26.98±1.03 |
16.48±0.85 |
|
25.63±1.13 |
17.18±1.12 |
|
Cu |
ns |
15.03±1.28 |
16.00±1.10 |
|
14.13±0.51 |
13.23±0.37 |
|
Fe |
ns |
109.13±3.93 |
119.50±5.30 |
|
108.25±1.80 |
116.50±5.55 |
|
Mn |
A* |
68.65±3.27 |
106.55±15.48 |
|
74.15±6.09 |
106.75±2.81 |
|
Zn |
ns |
20.18±1.37 |
22.30±1.84 |
|
21.85±1.79 |
21.50±1.10 |
|
|
|
|
|
|
|
|
2002 |
Al |
ns |
54.58±5.81 |
57.75±3.99 |
|
57.78±9.30 |
61.38±8.88 |
|
B |
A** |
29.78±1.33 |
16.93±0.41 |
|
28.38±0.93 |
16.28±2.13 |
|
Cu |
PM* |
346.00±80.05 |
346.25±39.33 |
|
15.65±1.38 |
20.25±1.25 |
|
Fe |
ns |
109.50±2.90 |
116.40±7.76 |
|
104.55±7.59 |
118.33±11.40 |
|
Mn |
A* |
103.98±12.39 |
165.50±18.23 |
|
111.55±10.36 |
146.25±13.42 |
|
Zn |
PM* |
22.20±1.39 |
21.75±1.01 |
|
33.05±5.56 |
31.03±2.40 |
|
|
|
|
|
|
|
|
2003 |
Al |
PM* |
30.63±1.36 |
37.98±3.46 |
|
43.30±2.39 |
35.33±1.93 |
|
B |
A** |
20.58±0.25 |
13.20±0.29 |
|
19.33±0.39 |
12.75±0.57 |
|
Cu |
ns |
11.96±1.38 |
13.50±0.18 |
|
11.35±0.82 |
12.78±0.92 |
|
Fe |
A* |
80.00±2.17 |
82.78±2.43 |
|
81.85±1.49 |
88.48±2.67 |
|
Mn |
A**, PM** |
59.43±4.22 |
132.50±10.10 |
|
128.50±9.08 |
182.25±16.73 |
|
Zn |
PM* |
17.00±0.80 |
16.90±0.86 |
|
21.38±1.19 |
22.43±2.26 |
Table 6. Multiple regression analyses for effects of mineral concentrations of potato leaves on the densities of different Colorado potato beetle life stages. Nutrients that were tested in the study but are not listed in this table did not have a statistically significant effect on beetle densities (P>0.05).
|
Parameter Estimates |
|
Model ANOVA |
|
Adjusted R2 |
||||||
|
Nutrient |
b |
St. Error |
t |
P |
|
df |
F |
P |
|
|
Overwintered adults |
|
|
|
|
|
|
|
|
|
||
|
K |
-11.91 |
4.43 |
-2.69 |
0.0083 |
|
11, 116 |
8.7 |
<0.0001 |
|
0.40 |
|
P |
-266.07 |
88.68 |
-3.00 |
0.0033 |
|
|
|
|
|
|
|
Zn |
0.25 |
0.09 |
2.70 |
0.0080 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Egg masses |
|
|
|
|
|
|
|
|
|
|
|
|
K |
-17.66 |
4.01 |
-4.41 |
<0.0001 |
|
11, 116 |
13.05 |
<0.0001 |
|
0.51 |
|
P |
-237.74 |
80.20 |
-2.96 |
0.0037 |
|
|
|
|
|
|
|
B |
-1.17 |
0.55 |
-2.12 |
0.0360 |
|
|
|
|
|
|
|
Zn |
0.33 |
0.08 |
3.93 |
0.0001 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Small larvae |
|
|
|
|
|
|
|
|
|
|
|
|
K |
11.30 |
4.39 |
2.57 |
0.0114 |
|
11, 116 |
9.1 |
<0.0001 |
|
0.41 |
|
B |
-3.15 |
0.60 |
-5.21 |
<0.0001 |
|
|
|
|
|
|
|
Zn |
0.38 |
0.09 |
4.21 |
<0.0001 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Large larvae |
|
|
|
|
|
|
|
|
|
|
|
|
P |
-217.06 |
75.39 |
-2.88 |
0.0048 |
|
11, 116 |
16.17 |
<0.0001 |
|
0.57 |
|
Al |
0.19 |
0.09 |
2.11 |
0.0374 |
|
|
|
|
|
|
|
B |
-1.64 |
0.52 |
-3.17 |
0.0019 |
|
|
|
|
|
|
|
Fe |
-0.22 |
0.11 |
-1.99 |
0.0490 |
|
|
|
|
|
|
|
Zn |
0.33 |
0.08 |
4.22 |
<0.0001 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Summer adults |
|
|
|
|
|
|
|
|
|
|
|
|
N |
-0.25 |
0.12 |
-2.11 |
0.0370 |
|
11, 116 |
12.61 |
<0.0001 |
|
0.50 |
|
B |
-2.23 |
0.56 |
-4.01 |
0.0001 |
|
|
|
|
|
|
|
Cu |
-0.30 |
0.10 |
-2.99 |
0.0034 |
|
|
|
|
|
|
|
Zn |
0.32 |
0.08 |
3.83 |
0.0002 |
|
|
|
|
|
|
Discussion
Substantially lower Colorado potato beetle populations were detected in the plots with manure-amended soil. Those results parallel the findings by Phelan et al. (1995, 1996) and Phelan (1997), who reported a substantial decrease in European corn borer (Ostrinia nubilalis [Hübner]) damage for plants grown in organically managed soil. Soil amendment was the major factor affecting beetle populations in our study. Neither crop rotation nor pest management approach appeared to be particularly important. Their interactions with soil amendment were also insignificant. The only exception was a significant interaction between pest management approach and soil amendment in 2002. However, it was observed during a single season only, and most likely did not have much biological meaning.
In contrast to beetle abundance, plant height and canopy cover were largely comparable between the plots receiving manure and synthetic fertilizer. Furthermore, tuber yields were higher in the manure-amended plots. Therefore, it is possible to conclude that lower beetle densities observed in those plots were not caused by smaller size or poor vigor of host plants. Similarly, Phelan et al. (1995) ruled out plant productivity as a factor determining European corn borer preference towards corn plants grown in the organically amended soils.
In direct accordance with the mineral balance hypothesis, there was a dramatic dissimilarity in the mineral composition of potato leaves collected from manure- and synthetic fertilizer-amended plots. Overall, differences were detected in concentrations of nitrogen, calcium, magnesium, phosphorus, aluminum, boron, copper, iron, manganese, and zinc. Except for phosphorus and boron, those were the same minerals that differed in the foliage of corn plants grown in organically and conventionally amended soils (Phelan et al., 1996). Crop rotation did not have consistent effects on the mineral content of potato leaves. Pest management approach had more of an effect, but it was still weaker and more sporadic than that of the soil amendment. Because EBDC fungicides applied in conventional plots contained manganese and zinc, and biorational IPM plots received copper hydroxide as part of an earlier study in 1991-98 (Gallandt et al., 1998), pest management influences on mineral concentrations most likely could be attributed to fungicide treatments. There was generally little interaction between the soil amendment and either crop rotation or pest management approach.
Mineral concentrations in potato leaves were responsible for 40-57% of the variation in the Colorado potato beetle populations observed among the experimental plots over the entire duration of the study. Zinc had a consistently positive effect on beetle populations, but rarely was significantly different between manure-amended and synthetic fertilizer-treated plots. Boron had a strong negative effect on all beetle stages except for the overwintered adults. Also, concentrations of this element were usually about two-fold higher in the plants grown on manure-amended soil. Beanland et al. (2003) observed increased developmental performance of the soybean looper (Pseudoplusia includens [Walker]), the Mexican bean beetle (Epilachna varivestis Mulsant), and the velvetbean caterpillar (Ancarsia gemmatalis Hübner) on soybean plants grown on hydroponic solutions that lacked boron. Therefore, it is possible that lower Colorado potato beetle densities observed in manure-amended plots were at least partially determined by the elevated boron concentrations.
Decrease in the beetle densities in manure-amended plots and its relation with the mineral composition of potato leaves strongly suggest that manure-amended soil had a buffering capability to maintain favorable nutrient balance in plants. However, it is important to remember that manure-amended plots were receiving organic amendments for eight years before the beginning of the present study. Therefore, it was impossible to distinguish between the immediate effect of manure amendments and long-term effect of soil management history. Phelan et al. (1995) demonstrated that multi-year organic management might be required for soils to acquire their buffering capacity.
Certainly, the level of pest suppression observed in the present study was hardly comparable to that provided by synthetic insecticides. Nevertheless, the decrease of the Colorado potato beetle populations shows yet another benefit of organic soil amendments for crop fertilization. This should be one of the considerations when designing fully integrated crop management systems.
Acknowledgments
We thank David Lambert and Eric Gallandt for their help with disease and weed management, respectively. We also thank Erwin Ganga, Paul Ocaya, Gary Sewell, Jonathan Sisson, and many others for general technical assistance. P. Larry Phelan provided helpful comments on the earlier version of this manuscript. This work was supported in part by USDA-CSREES-IFAFS No. 2001-52101-11308, the University of Maine/CSREES Hatch Project No. ME08912-02, and the Maine Potato Board. This is Publication No. 2772 of the Maine Agricultural and Forest Experiment Station.
References
Beanland, L., Phelan, P.L., Salminen, S., 2003. Micronutrient interactions on soybean growth and the developmental performance of three insect herbivores. Environ. Entomol. 32, 641-651.
Busch, J.W., Phelan, P.L., 1999. Mixture models of soybean growth and herbivore performance in response to nitrogen-sulphur-phosphorous nutrient interactions. Ecol. Entomol. 24, 132-145.
Casagrande, R.A., 1987. The Colorado potato beetle: 125 years of mismanagement. Bull. Entomol. Soc. Am. 33, 142-150.
Clancy, K.M., 1992. Response of western spruce budworm (Lepidoptera: Tortricidae) to increased nitrogen in artificial diets. Environ. Entomol. 21, 331-344.
Clancy, K.M., King, R.M., 1993. Defining the western spruce budworm’s nutritional niche with response surface methodology. Ecology 74, 442-454.
Clancy, K.M., Wagner, M.R., Tinus, R.W., 1988. Variation in host foliage nutrient concentrations in relation to western spruce budworm herbivory. Can. J. Forest Res. 18, 530-539.
Conover, W.J., Iman, R.L., 1981. Rank transformations as a bridge between parametric and nonparametric statistics. Am. Stat. 35, 124-129.
Delate, K., Friedrich, H., Lawson, V., 2003. Organic pepper production systems using compost and cover crops. Biol. Agric. Hort. 21, 131-150.
Drummond, F.A., Groden, E., 1996. Insect Pests and Natural Enemies. In: Marra, M.C. (Ed.), The Ecology, Economics, and Management of Potato Cropping Systems: A Report of the First Four Years of the Maine Potato Ecosystem Project. Maine Agric. and Forest Experiment Station Bull. 843, pp 80-118.
Dwyer, J.D., Johnson, S.B., Kleinhenz, M.D., Plissey, E.S., 1997. Maine Potato Pest Control Guide. Univ. Maine Coop. Ext. Bull. #2002.
Eigenbrode, S.D., Pimentel, D., 1988. Effects of manure and chemical fertilizers on insect pest populations on collards. Agric. Ecosystems Environ. 20, 109-125.
Feber, R. E., Firbank, L. G., Johnson, P. J., McDonald, D. W., 1997. The effects of organic farming on pest and non-pest butterfly abundance. Agric. Ecosystems Environ. 64, 133-139.
Gallandt, E.R., Mallory, E.B., Alford, A.R., Drummond, F.A., Groden, E., Liebman, M., Marra, M.C., McBurnie, J.C., Porter, G.A., 1998. Comparison of alternative pest and soil management strategies for Maine potato production systems. Am. J. Altern. Agric. 13, 146-161.
Huynh, H., Feldt. L.S., 1970. Conditions under which mean square ratios in repeated measurement designs have exact F-distributions. J. Am. Stat. Assoc. 65, 1582-1589.
Letourneau, D. K., Goldstein, B., 2001. Pest damage and arthropod community structure in organic vs. conventional tomato production in California. J. Appl. Ecol. 38, 557-570.
Oelhaf, R.C., 1978. Organic Farming: Economic and Ecological Comparisons with Conventional Methods. Allanheld, Osmun, Montclair, N.J.
Phelan, P. L., 1997. Soil-management history and the role of plant mineral balance as a determinant of maize susceptibility to the European corn borer. Biol. Agric. Hortic. 15, 25-34.
Phelan, L. P., Mason, J. F., Stinner, B.R., 1995. Soil-fertility management and host preference by European corn borer, Ostrinia nubilalis (Hübner), on Zea mays L.: a comparison of organic and conventional chemical farming. Agric. Ecosystems Environ. 56, 1-8.
Phelan, L.P., Norris, K.H., Mason, J. F., 1996. Soil management history and host preference by Ostrinia nubilalis: evidence for plant mineral balance mediating insect-plant interactions. Environ. Entomol. 25, 1329-1336.
Porter, G.A., 1996. Appendix A – General Methods. In: Marra, M.C. (Ed.), The Ecology, Economics, and Management of Potato Cropping Systems: A Report of the First Four Years of the Maine Potato Ecosystem Project. Maine Agric. and Forest Experiment Station Bull. 843, pp 161-174.
Porter, G.A., McBurnie, J.C., 1996. Crop and Soil Research. In: Marra, M.C. (Ed.), The Ecology, Economics, and Management of Potato Cropping Systems: A Report of the First Four Years of the Maine Potato Ecosystem Project. Maine Agric. and Forest Experiment Station Bull. 843, pp 8-62.
SAS Institute, Inc., 1999. SAS OnLine Doc, Version 8. Cary, North Carolina.
Weber, D.C., Ferro, D.N., 1994. Colorado potato beetle: diverse life history poses challenge to management. In: Zehnder, G. W., Jansson, R. K., Powelson, M. L. and Raman, K. V. (Eds.), Advances in Potato Pest Biology and Management. APS Press, St. Paul, pp 54-70.