

For a more scientific review, please refer to
Alyokhin, A. 2009. Colorado potato beetle management on potatoes: current challenges and future prospects. In: Tennant P, Benkeblia N (Eds) Potato II. Fruit, Vegetable and Cereal Science and Biotechnology 3 (Special Issue 1): 10-19.
Available free of charge with permission of the publisher, Global Science Books Ltd.
Introduction
Colorado potato beetle (Leptinotarsa decemlineata [Say]) is the most important insect defoliator of potatoes. It also causes significant damage to tomato and eggplant. One beetle consumes approximately 40 cm2 of potato leaves at a larval stage, and up to additional 9.65 cm2 of foliage per day as an adult (Ferro et al., 1985). In addition to impressive feeding rates, Colorado potato beetle is also characterized by high fecundity, with one female laying 300-800 eggs (Harcourt, 1971). Furthermore, the beetle has a remarkable ability to develop resistance to virtually every chemical that has ever been used against it.
Distribution
Since Colorado potato beetle shifted from its original wild hosts in southwestern North America, it has spread throughout the rest of the continent and has invaded Europe and Asia. Currently its distribution covers about 8 million km2 in North America (Hsiao, 1985) and about 6 million km2 in Europe and Asia (Jolivet, 1991). It has appeared recently in western China and Iran. Potentially the Colorado potato beetle can occupy much larger areas in China and Asia Minor, spread to Korea, Japan, Russian Siberia, certain areas of the Indian subcontinent, parts of North Africa, and the temperate Southern Hemisphere (Vlasova, 1978; Worner, 1988; Jolivet, 1991).
Life History
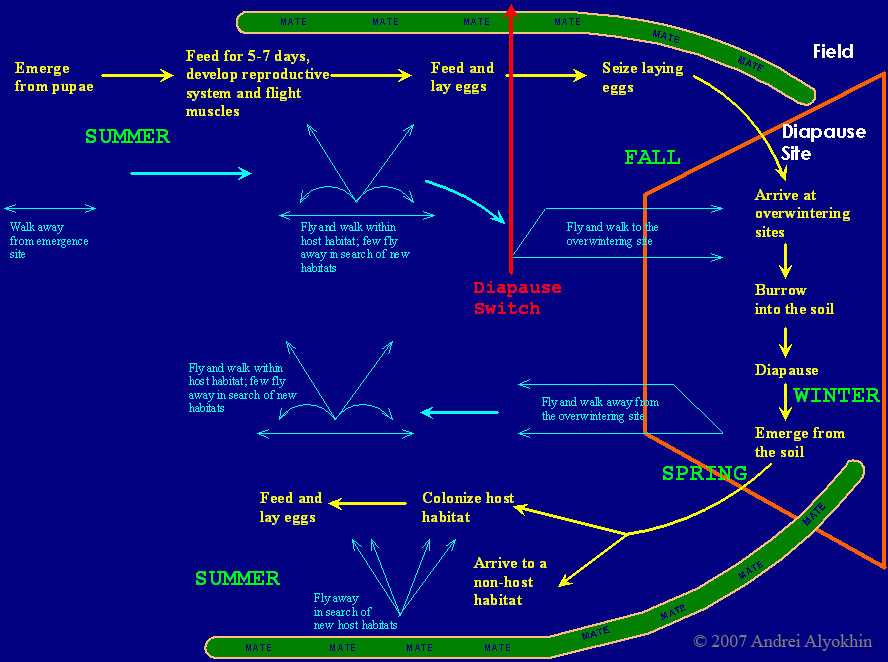
The Colorado potato beetle has a complicated and diverse life history. The beetles overwinter in the soil as adults, with the majority aggregating in woody areas adjacent to fields where they have spent the previous summer (Weber and Ferro, 1993). The emergence of post-diapause beetles is more or less synchronized with potatoes. If fields are not rotated, they are colonized by overwintered adults that walk to the field from their overwintering sites or emerge from the soil within the field (Voss and Ferro, 1990). If fields are rotated, the beetles are able to fly up to several kilometers to find a new host habitat (Ferro et al., 1991; 1999). Once they have colonized the field, the overwintered beetles first feed and then oviposit within 5-6 days depending on temperature (Ferro et al., 1985; Ferro et al., 1991).
Eggs are usually laid on the underside of potato leaves. Upon hatching, larvae may move over short distances within potato canopy and start feeding within 24 hours of eclosion. Development from the time of oviposition to adult eclosion for pupae takes between 14-56 days (de Wilde, 1948; Walgenback and Wyman, 1984; Logan et al., 1985; Ferro et al., 1985). The optimal temperatures range between 25-32ºC and appear to differ among populations of different geographic origins. The larvae are capable of behavioral thermoregulation via moving within plant canopies (May, 1981; Lactin and Holliday, 1994), thus optimizing their body temperature compared to the ambient temperature. Pupation takes place in the soil near the plants where the larval development has been completed.
Diapause is facultative, and the beetles can have between one and three overlapping generations per year. It takes a few days for the newly emerged adults to develop their reproductive system and flight muscles (Alyokhin and Ferro, 1999). After development has been completed, the beetles mate and start laying eggs. The reproduction continues until diapause is induced by the short-day photoperiod, then the beetles migrate to overwintering sites (mainly by flying), and enter the soil to diapause. Those beetles that emerge under short-day photoperiod do not develop their reproductive system and flight muscles that season. They feed actively for several weeks and then either walk to the overwintering sites or burrow into the soil directly in the field (Voss, 1989).
Colorado potato beetle's diverse and flexible life history is well-suited to unstable agricultural environments, and makes it a complex and challenging pest to control. Flight migrations closely connected with diapause, feeding and reproduction allow the Colorado potato beetle to employ "bet-hedging" reproductive strategies, distributing its offspring in both space (within and between fields) and time (within and between years). Such strategies minimize the risk of catastrophic losses of offspring, otherwise quite possible in unstable agricultural ecosystems (Solbreck, 1978; Voss and Ferro, 1990).
Chemical Control and Insecticide Resistance
Since 1864, hundreds of compounds were tested against the Colorado potato beetle (Gauthier et al. 1981), and insecticides still remain the foundation of the Colorado potato beetle control on commercial potato farms. Currently, more than 30 active ingredients are registered for use against this pest in the United States. Insecticide efficiency and availability, however, vary from area to area. The same is true about pesticide regulations. Therefore, people considering using chemicals to control beetles should contact a local extension agent or other qualified professional.
An important thing to keep in mind is that Colorado potato beetle has a legendary ability to develop resistance to a wide range of pesticides used for its control. High predisposition to resistance development seems to be an inherent characteristic of this species. It is probably caused, in large part, by the coevolution of the beetle and its host plants in the family Solanaceae, which have high concentrations of toxins, namely glycoalkaloids (Ferro, 1993). The first instance of Colorado potato beetle resistance to synthetic organic pesticides was noted for DDT in 1952 (Quinton, 1955). Resistance to dieldrin was reported in 1958, followed by resistance to other chlorinated hydrocarbons (Hofmaster et al., 1967). In subsequent years the beetle has developed resistance to numerous organophosphates and carbamates (Forgash, 1985). Presently it is resistant to a wide range of insecticides, including the arsenicals, organochlorines, carbamates, organophosphates, and pyrethroids. Resistance crisis was temporarily abated with the introduction of highly effective neonicotinoid insecticides. However, the first cases of beetle resistance to neonicotinoids have been already observed in several field populations (Alyokhin et al., 2006; 2007; Mota-Sanchez et al., 2006).
The major problem area is the Northeastern United States (Forgash, 1985); however, resistance has also been detected in Michigan (Ioannidis et al., 1991), Canada (Stewart et al., 1997), and Europe (Forgash, 1985; Boiteau, 1988). In some cases, a new insecticide failed after one year (e.g., endrin) or even during the first year of use (e.g., oxamyl) (Forgash, 1985). Resistance mechanisms are highly diverse even within a relatively narrow geographical area (Ioannidis et al., 1991). Furthermore, the beetles show cross-resistance to organophosphates and carbamates, and multiple resistance to organophosphates, carbamates, and pyrethroids (Ioannidis et al., 1991). In addition to the resistance to synthetic insecticides, the beetle has a capability to develop resistance to the Bacillus thuringiensis subsp. tenebrionis delta-endotoxin (Whalon et al., 1993; Rahardja and Whalon, 1995).
Cultural Control
Colorado potato beetle populations can be reduced through the use of relatively common cultural practices such as crop rotation, manipulation of planting time and crop varieties, use of mulches, cover and trap crops (Hough-Goldstein et al., 1993). Crop rotation for the Colorado potato beetle control had been first recommended as early as 1872 (Bethune, 1872), and since then proved to be a good control strategy not only for the beetle, but also for a number of potato pathogenes and weeds (Casagrande, 1987). At the rotated field, peak density of the beetle egg masses could be less than 10% of that of the non-rotated field (Lashomb and Ng, 1984). Wright (1984) reported that when potatoes were planted following a non-host grain crop (rye or wheat), early season Colorado potato beetle adult densities were reduced by 95.8%.
Late and early planting is aimed to suppress the second generation larval populations. Because summer-generation adults emerge later in the season on the late-planted crop, the short-day photoperiod stimulates reproductive diapause, largely eliminating the second-generation larval impact on the crop. Early planting also eliminates the second generation larvae, in this case because the crop is already being removed at the time of their emergence (Weber and Ferro, 1995).
Trap crops may be used to attract beetles away from the main crop. It has been shown to intercept both overwintered beetles colonizing a field in the spring (Weber and Ferro, 1995), as well as the beetles moving away from senescing potatoes late in the season (Hoy et al., 1996).
Another promising technique of Colorado potato beetle cultural control is mulch applications. Larval populations of the beetle were significantly reduced in straw- mulched plots of potato (Stoner, 1993) and eggplant (Stoner, 1997). A peak of the small (1st - 2nd instar) larval populations was observed 1 - 2 weeks later on the mulched potato fields than on the unmulched ones (Stoner, 1993). Furthermore, the mulch may increase the time required by the beetles to find potatoes (Ng and Lashomb, 1983), decrease the likelihood of flying beetles locating the potato plants (East, 1993), increase the proportion of beetles leaving the area by flight (Weber et al., 1994), and increase predation on eggs and larvae (Brust, 1994). Overall, a six - ten cm layer of wheat straw produced 2.5-5 fold decrease in potato defoliation (Zehnder and Hough-Goldstein, 1990; Brust, 1994).
Physical Control
In addition to cultural control, a number of physical control methods can be used to suppress Colorado potato beetle populations. One possible method involves digging plastic-lined trenches along a field border in order to intercept post-diapause Colorado potato beetles colonizing the crop in the spring. In a one-month period, during which the majority of the beetles emerge from the soil, 1 m of such a trench can capture as many as 1,000 beetles (Ferro, unpublished data). Up to 95% of captured beetles are normally retained in the ditch (Misener et al., 1993).
Another method is to manipulate beetle diapause habitat in an attempt to enhance its overwintering mortality. In the experiment of Milner et al. (1992), wheat straw mulch was applied to the overwintering sites in the fall, and then removed together with the layer of snow covering it in January. This procedure rapidly depressed soil temperatures, and led to a significantly lower beetle survival (approximately 7% at disturbed habitats vs. approximately 26% at the undisturbed habitats).
Still other methods of physical control include propane flamers (Pelletier et al., 1995; Khelifi et al., 2007) and tractor-mounted vacuum collectors (Boiteau et al., 1992; Lacasse et al., 1998). Combining these two techniques increases their overall efficiency (Khelifi et al. 2007), making control level comparable to that of some insecticide treatments (Laguë et al., 1999).
Biological Control
High fecundity usually allows Colorado potato beetle populations to withstand natural enemy pressure. Still, in the absence of insecticides natural enemies can sometimes reach densities capable of reducing Colorado potato beetle numbers below economically damaging levels (Ferro, 1985).
Beauveria bassiana (Hyphomycetes) is a pathogenic fungus that infects a wide range of insect species, including the Colorado potato beetle. It is probably the most widely used natural enemy of the Colorado potato beetle, with readily available commercial formulations that can be applied using a regular pesticide sprayer. Applications of B. bassiana have been shown to reduce beetle populations by up to 75% (Cantwell et al.,1986). However, control is usually less effective compared to chemical insecticides (Campbell et al., 1985; Hajek et al., 1987).
A number of predatory and parasitic arthropods attack the Colorado potato beetle (Hough-Goldstein et al., 1993). The lady beetle Coleomegilla maculata consumes eggs and small larvae (Groden et al., 1990; Hazzard et al., 1991), killing up to 37.8% of eggs for the first Colorado potato beetle generation and up to 58.1% of eggs for the second generation (Hazzard et al., 1991). Predaceous stink bugs Perillus bioculatus and Podisus maculiventris attack beetle larvae. Inundative releases of these predators suppressed beetle density by 62% (Biever and Chauvin, 1992), reduced defoliation by 86% (Hough-Goldstein and McPherson, 1996), and increased potato yields by 65% (Biever and Chauvin, 1992) over the untreated control. Adult ground beetles Lebia grandis feed on the Colorado potato beetle eggs and larvae, while larvae of the same species parasitize the Colorado potato beetle pupae (Weber et al., 2006). The parasitic wasp Edovum puttleri was found to parasitize 71-91% of Colorado potato beetle egg masses on eggplant, killing 67-79% of the eggs per mass (Lashomb et al., 1987). The level of parasitism is somewhat lower in potatoes, rarely exceeding 50% (Ruberson et al., 1991; Van Driesche et al., 1991). Performance of Edovum puttleri in the field can be further improved by the supplementary use of an artificial carbohydrate source (Idoine and Ferro, 1990).
Several species of generalist predators also occasionally feed on the Colorado potato beetle. Fourteen species of carabid beetles, three species of Coccinellidae, and a spider, Xysticus kochi, are known to feed on the Colorado potato beetle in the former Soviet Union (Sorokin, 1976). Eight species of Lebia and five other ground beetle species attack this pest in Mexico (Logan, 1990). Another ground beetle, Pterostichus chalcites, has been observed feeding on the Colorado potato beetle in Delaware (Heimpel and Hough- Goldstein, 1992). The daddy-long-legs Phalangium opilio preys on the beetle's eggs and small larvae (Drummond et al., 1990).
Plant Resistance
Although Colorado potato beetles are fully capable of completely wiping out potato crops, at low-to-moderate beetle densities potato plants are fairly tolerant to the inflicted defoliation. They can tolerate 30-40% defoliation during early growth stages, 10-60% defoliation during middle growth stages, and up to 100% defoliation late in the season without noticeable yield reduction (Hare, 1980; Cranshaw and Radcliffe, 1980; Ferro et al., 1983; Shields and Wyman, 1984; Zehnder and Evanylo, 1988). In the same time, currently there are no truly resistant cultivars. Conventional potato breeding is complicated by tetraploidy in S. tuberosum (Grafius and Douches 2008). Genetically modified potatoes expressing Bacillus thuringiensis delta-endotoxin that is toxic to the Colorado potato beetle were introduced in the U.S. in 1995, but then discontinued after only five years of use, in large part because of consumer concerns about genetically engineered foods.
Cultural practices may enhance plant ability to resist the Colorado potato beetle. Lower beetle densities have been recorded in plots receiving manure soil amendments in combination with reduced amounts of synthetic fertilizers compared to plots receiving full rates of synthetic fertilizers, but no manure (Alyokhin et al., 2005). No reduction in plant vigor in the absence of synthetic fertilizers was observed in that study. Subsequent field-cage and laboratory experiments (Alyokhin and Atlihan, 2005) confirmed that potato plants grown in manure-amended soil were indeed inferior Colorado potato beetle hosts compared to plants grown in synthetically fertilized soil.
Integrated Pest Management
The secret of Colorado potato beetle’s success as a pest is its diverse and flexible life history coupled with a remarkable adaptability. Therefore, to be successful in our control efforts we also need to be diverse and flexible in our approaches, as well as adaptable to ever-changing circumstances. Mindless reliance on a single tactic is doomed to fail, no matter how fundamentally sound this tactic is. The only sustainable way to manage this insect is integration of multiple control techniques based on a scientifically sound understanding of its biology.
Control in Home Gardens
In addition to creating problems in commercial production, the Colorado potato beetle is also a concern for home gardeners. When garden is limited to a few potato, tomato, or eggplant plants, hand-picking overwintered adults and egg masses early in the season is the simplest management approach. Most damage is done by larvae, so removing their parents and unhatched eggs should provide fairly good protection of the plants later in the season. It is no more time-consuming than other gardening practices, does not require expensive purchased inputs, and environmentally friendly. It can also be a relaxing and somewhat therapeutic experience – after all, from the biological point of view we have evolved to be hunters and gatherers, not computer programmers or hedge fund managers. The picking should be done for several weeks because overwintered beetles exit diapause and colonize host plants over approximately one-month time window.
References Cited
Alyokhin, A.V. and D.N. Ferro. 1999. Reproduction and dispersal of summer-generation Colorado potato beetle (Coleoptera: Chrysomelidae). Environ. Entomol. 28: 425-430.
Alyokhin, A. and R. Atlihan. 2005. Reduced fitness of the Colorado potato beetle (Coleoptera: Chrysomelidae) on potato plants grown in manure-amended soil. Environ. Entomol. 34: 963-968.
Alyokhin, A., G. Porter, E. Groden, and F. Drummond. 2005. Colorado potato beetle response to soil amendments: a case in support of the mineral balance hypothesis? Agric. Ecosyst. Environ 109: 234-244.
Alyokhin, A., G. Dively, M. Patterson, D. Rogers, M. Mahoney, and J. Wollam. 2006. Susceptibility of imidacloprid-resistant Colorado potato beetles to non-neonicotinoid insecticides in the laboratory and field trials. Am. J. Potato Res. 83: 485-494.
Alyokhin, A., G. Dively, M. Patterson, C. Castaldo, D. Rogers, M. Mahoney, and J. Wollam. 2007. Resistance and cross-resistance to imidacloprid and thiamethoxam in the Colorado potato beetle. Pest Manage. Sci. 63: 32-41.
Bethune, C.J.S. 1872. Report of the Entomological Society of Ontario for the year 1871. Hunter, Ross, Toronto.
Boiteau, G. 1988. Control of the Colorado potato beetle, Leptinotarsa decemlineata (Say): learning from the Soviet experience. Bull. Entomol. Soc. Can. 20: 9-14.
Boiteau, G., G.C. Misener, R.P. Singh, and G. Bernard. 1992. Evaluation of a vacuum collector for insect pest control in potato. Am. Potato J. 69: 157-166.
Brust, G.E. 1994. Natural enemies in straw-mulch reduce Colorado potato beetle populations and damage in potato. Biol. Control 4: 163-169.
Biever, K.D. and R.L. Chauvin. 1992. Suppression of the Colorado potato beetle (Coleoptera: Chrysomelidae) with augmentative releases of predaceous stinkbugs (Hemiptera: Pentatomidae). J. Econ. Entomol. 85: 720-726.
Campbell, R.K., T.E. Anderson, M. Semel, and D.W. Roberts. 1985. Management of the Colorado potato beetle using the entomogenous fungus Beauveria bassiana. Am. Potato J. 62: 29-37.
Cantwell, G.E., W.W. Cantelo, and R.F.W. Schroder. 1986. Effect of Beauveria bassiana on underground stages of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Great Lakes Entomologist 19, 81- 84.
Casagrande, R.A. 1987. The Colorado potato beetle: 125 years of mismanagement. Bull. Entomol. Soc. Am. 33: 142-150.
Cranshaw, W.S. and E.B. Radcliffe. 1980. Effect of defoliation on yield of potatoes. J. Econ. Entomol 73: 131-134.
de Wilde J. 1948. Developpement embryonnaire et postembryonnaire dy doryphore (Leptinotarsa decemlineata Say) en function de la temperature. In: Proceedings 8th International Congress of Entomology, Stockholm, Sweden, pp. 310-321.
Drummond, F., Y. Suhaya and E. Groden. 1990. Predation on the Colorado potato beetle (Coleoptera: Chrysomelidae) by Phalangium opilio (Opiliones: Phalangidae). J. Econ. Entomol. 83: 772-778.
East, D.A. 1993. Colonization of potato fields by post-diapause Colorado potato beetles, Leptinotarsa decemlineata (Say). Ph.D. Dissertation, Ohio State University.
Ferro, D.N., B.J. Morzuch, and D. Margolies. 1983. Crop loss assessment of the Colorado potato beetle (Coleoptera: Chrysomelidae) on potatoes in western Massachusetts. J. Econ. Entomol. 76: 349-356.
Ferro, D.N. 1985. Pest status and control strategies of the Colorado potato beetle. In Ferro, D.N. and R.H. Voss (eds.) Proceedings of the Symposium on the Colorado potato beetle, XVII Inernational Congress of Entomology.
Ferro, D.N. 1993. Potential for resistance to Bacillus thuringiensis: Colorado potato beetle (Coleoptera: Chrysomelidae) - a model system. Am. Entomol. 39: 38-44.
Ferro, D.N., J.A. Logan, R.H. Voss, and J.S. Elkinton. 1985. Colorado potato beetle (Coleoptera: Chrysomelidae) temperature-dependent growth and feeding rates. Environ. Entomol. 14: 343-348.
Ferro, D.N., A.F. Tuttle, D.C. Weber. 1991. Ovipositional and flight behavior of overwintered Colorado potato beetle (Coleoptera: Chrysomelidae). Environ. Entomol. 20: 1309-1314.
Ferro, D. N., A. V. Alyokhin, and D. B. Tobin. 1999. Reproductive status and flight activity of the overwintered Colorado potato beetle. Entomol. Exp. Appl. 91: 443-448.
Forgash, A.G. 1985. Insecticide resistance in the Colorado potato beetle. In Ferro, D.N. and R.H. Voss (eds.) Proceedings of the Symposium on the Colorado potato beetle, XVII International Congress of Entomology.
Gauthier, N.L., R.N. Hofmaster, and M. Semel. 1981. History of Colorado potato beetle control. In: JH Lashomb and R Casagrande (eds). Advances in Potato Pest Management. Hutchinson Ross Publishing Co., Stroudsburg, PA, pp. 13-33.
Grafius, E.J. and S. Douches. 2008. The present and future role of insect-resistant genetically modified potato cultivars in potato IPM. In: Romeis J, Shelton AM, George G Kennedy (Eds) Integration of Insect-Resistant GM crops within IPM Programs, Springer, New York, pp 195-222.
Groden, E., F.A. Drummond, R.A. Casagrande and D.L. Haynes. 1990. Coleomegilla maculata (Coleoptera: Coccinellidae): its predation upon the Colorado potato beetle (Coleoptera: Chrysomelidae) and its incidence in potatoes and surrounding crops. J. Econ. Entomol. 83: 1306-1315.
Hajek, A.E., R.S. Soper, D.W. Roberts, T.E. Anderson, K.D. Biever, D.N. Ferro, R.A. Lebrun, and R.H. Storch. 1987. Foliar applications of Beauveria bassiana (Balsamo) Vuillemin for control of the Colorado potato beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae): an overview of pilot test results from the northern United States. Can. Entomol. 119: 959-974.
Harcourt, D.G. 1971. Population dynamics of Leptinotarsa decemlineata (Say) in eastern Ontario. III. Major population processes. Can. Entomol. 103: 1049-1061.
Hare, J.D. 1980. Impact of defoliation by the Colorado potato beetle Leptinotarsa decemlineata on potato yields. J. .Econ. Entomol 73: 369-373.
Hazzard, R.V., D.N. Ferro, R.G. Van Driesche, and A.F. Tuttle. 1991. Mortality of eggs of Colorado potato beetle (Coleoptera: Chrysomelidae) from predation by Coleomegilla maculata (Coleoptera: Coccinellidae). Environ. Entomol. 20: 841-848.
Heimpel, G.E. and J.A. Hough-Goldstein. 1992. A survey of arthropod predators of Leptinotarsa decemlineata (Say) in Delaware potato fields. J. Agric. Entomol. 9: 137-142.
Hofmaster, R.N., R.L. Waterfield, and J.C. Boyd. 1967. Insecticides applied to the soil for control of eight species of insects on Irish potatos in Virginia. J. Econ. Entomol. 60: 1311-1318.
Hough-Goldstein, J.A., G.E. Heimpel, H.E. Bechmann, and C.E. Mason. 1993. Arthropod natural enemies of the Colorado potato beetle. Crop Protection 12: 324-334.
Hough-Goldstein, J.A. and D. McPherson. 1996. Comparison of Perillus bioculatus and Podisus maculiventris (Hemiptera: Pentatomidae) as potential control agents of the Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 89: 1116-1123.
Hoy, C.W., J.A. Wyman, T.T. Vaughn, D.A. East, and P. Kaufman. 1996. Food, ground cover, and Colorado potato beetle (Coleoptera: Chrysomelidae) dispersal in late summer. J. Econ. Entomol. 89: 963-969.
Hsiao, T.H. 1985. Ecophysiological and genetic aspects of geographic variations of the Colorado potato beetle. In Ferro, D.N., Voss, R.H. (eds.) Proceedings of the Symposium on the Colorado potato beetle, XVII Inernational Congress of Entomology.
Idoine, K. and D.N. Ferro. 1990. Persistence of Edovum puttleri (Hymenoptera: Eulophidae) on potato plants and parasitism of Leptinotarsa decemlineata (Coleoptera: Chrysomelidae): effects of resource availability and weather. Environ. Entomol. 19: 1732-1737.
Ioannidis, P.M., E. Grafius, M.E. Whalon. 1991. Patterns of insecticide resistance to azinphosmethyl, carbofuran, and permethrin in the Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 84: 1417-1423.
Jolivet, P. 1991. The Colorado beetle menaces Asia (Leptinotarsa decemlineata Say) (Coleoptera: Chrysomelidae). L'Entomologiste. 47: 29-48.
Karalus, W. 1994. Control of potato beetles in organic farming. Kartoffelbau 145: 170-173.
Lacasse B., C. Laguë, M. Khelifi, and P.-M. Roy. 1998. Field evaluation of pneumatic control of Colorado potato beetle. Can. Agric. Engineering 40: 273-280.
Lactin, D.J. and N.J. Holliday . 1994. Behavioral responses of Colorado potato beetle larvae to combinations of temperature and insolation, under field conditions. Entomol. Exp. Appl. 72: 255-263.
Laguë C, M. Khelifi, J. Gill, and B. Lacasse. 1999. Pneumatic and thermal control of Colorado potato beetle. Can. Agric. Engineering 41: 053-057
Lashomb, J.H. and Y.S. Ng. 1984. Colonization by the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) in rotated and non- rotated potato fields. Environ. Entomol. 13: 1352-1356.
Lashomb, J.H., Y.S. Ng, R.K. Jansson and R. Bullock. 1987. Edovum puttleri (Hymenoptera: Eulophidae), an egg parasitoid of Colorado potato beetle (Coleoptera: Chrysomelidae): development and parasitism on eggplant. J. Econ. Entomol. 80: 65-68.
Logan, P.A. 1990. Summary of biological control activities. University of Rhode Island, 1989-1990. Natural Enemy News 2: 10.
Logan, P.A., R.A. Casagrande, H.H. Faubert, and F.A. Drummond. 1985. Temperature-dependent development and feeding of immature Colorado potato beetles, Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae). Environ. Entomol. 14:275-283.
May, M.L. 1981. Role of body temperature and thermoregulation in the biology of the Colorado potato beetle. In: JH Lashomb and R Casagrande (eds). Advances in Potato Pest Management. Hutchinson Ross Publishing Co., Stroudsburg, PA, pp. 86-104.
Milner, M., K.J.S. Kung, J.A. Wyman, J. Feldman, and E. Nordheim. 1992. Enhancing overwintering mortality of Colorado potato beetle (Coleoptera: Chrysomelidae) by manipulating the temperature of its diapause habitat. J. Econ. Entomol. 85: 1701- 1708.
Misener, G.C., G. Boiteau, and L.P. McMillan. 1993. A plastic-lining trenching device for the control of Colorado potato beetle: Beetle Excluder. Am. Potato J. 70: 903- 908.
Mota-Sanchez D, R.M. Hollingworth, E.J. Grafius, and D.D. Moyer. 2006. Resistance and cross-resistance to neonicotinoid insecticides and spinosad in the Colorado potato beetle, Leptinotarsa decemlineata (Say) (Coleoptera : Chrysomelidae). Pest Manage. Sci. 62: 30-37.
Ng, Y.S. and J. Lashomb. 1983. Orientation by the Colorado potato beetle (Leptinotarsa decemlineata Say). Anim. Behav. 31: 617-619.
Quinton, R.J. 1955. DDT-resistant Colorado potato beetles? Proc. North Centr. Ent. Soc. Am. 9: 94-95.
Pelletier, Y., C.D. McLeod, and G. Bernard. 1995. Description of sublethal injuries caused to the Colorado potato beetle (Coleoptera: Chrysomelidae) by propane flamer treatment. J. Econom. Entomol. 88: 1203-1205.
Rahardja, U. and M.E. Whalon. 1995. Inheritance of resistance to Bacillus thuringiensis subsp. tenebrionis CryIIIA delta-endotoxin in Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 88: 21-26.
Ruberson, J.R., M.J. Tauber, C.A. Tauber, B. Gollands. 1991. Parasitization by Edovum puttleri (Hymenoptera: Eulophidae) in relation to host density in the field. Ecol. Entomol. 16: 81-89.
Shields, E.J., and J.A. Wyman. 1984. Effect of defoliation at specific growth stages on potato yields. J. Econ. Entomol. 77: 1194-1199.
Solbreck, C. 1978. Migration, diapause, and direct development as alternative life histories in a seed bug, Neacoryphus bicrucis. In Dingle, H. (ed.) Evolution of insect migration and diapause. Springer, New York.
Sorokin, N.S. 1976. The Colorado potato beetle (Leptinotarsa decemlineata Say) and its entomophages in the Rostov Region. Biull. Vses. Nauchn. Issled. Inst. Zashch. Rast. 37: 22-27.
Stewart, J.G., G.G. Kennedy, and A.V. Sturz. 1997. Incidence of insecticide resistance in populations of Colorado potato beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae), on Prince Edward Island. Can. Entomol. 129: 21- 26.
Stoner, K.A. 1993. Effects of straw and leaf mulches and sprinkle irrigation on the abundance of Colorado potato beetle (Coleoptera: Chrysomelidae) on potato in Connecticut. J. Entomol. Sci. 28: 393-403.
Stoner, K.A. 1997. Influence of mulches on the colonization by adults and survival of larvae of the Colorado potato beetle (Coleoptera: Chrysomelidae) in eggplant. J. Entomol. Sci. 32: 7-16.
Van Driesche, R.G., D.N. Ferro, E. Carey, and M. Maher. 1991. Assessing augmentative releases of parasitoids using the "recruitment method", with reference to Edovum puttleri, a parasitoid of the Colorado potato beetle (Coleoptera: Chrysomelidae). Entomophaga 36: 193-204.
Vlasova, V.A. 1978. A prediction of the distribution of Colorado beetle in the Asiatic territory of the USSR. Zaschita Rastenii 6: 44-45.
Voss, R.H. 1989. Population dynamics of the Colorado potato beetle, (Leptinotarsa decemlineata) (Say) (Coleoptera: Chrysomelidae), in western Massachusetts, with particular emphasis on migration and dispersal processes. Ph.D. Thesis, University of Massachusetts, Amherst.
Voss, R.H., D.N. Ferro. 1990. Phenology of flight and walking by Colorado potato beetle (Coleoptera: Chrysomelidae) adults in western Massachusetts. Environ. Entomol. 19: 117-122.
Walgenbach, J.F. and J.A. Wyman. 1984. Colorado potato beetle (Coleoptera: Chrysomelidae) development in relation to temperature in Wisconsin. Ann. Entomol. Soc. Am. 77: 604-609.
Weber, D.C. and D.N. Ferro. 1993. Distribution of overwintering Colorado potato beetle in and near Massachusetts potato fields. Entomol. Exp. Appl. 66: 191-196.
Weber, D.C., D.N. Ferro, J. Buonaccorsi, and R.V. Hazzard. 1994. Disrupting spring colonization of Colorado potato beetle to nonrotated potato fields. Ent. Exp. Appl. 73: 39-50.
Weber, D.C. and D.N. Ferro, 1995. Colorado potato beetle: diverse life history holds keys to management. In G.W. Zehnder, R.K. Jansson, M.L. Powelson and K.V. Raman (eds.), Advances in Potato Pest Biology and Management. American Phytopathological Society Press, St. Paul, MN.
Weber, D.C., D.L. Rowley, M.H. Greenstone, and M.M. Athanas. 2006. Prey preference and host suitability of the predatory and parasitoid carabid beetle, Lebia grandis, for several species of Leptinotarsa beetles. 14pp. Journal of Insect Science 6:09, available online: insectscience.org/6.09
Whalon, M.E., D.L. Miller, R.M. Hollingworth, E.J. Grafius, and J.R. Miller. 1993. Selection of a Colorado potato beetle (Coleoptera: Chrysomelidae) strain resistant to Bacillus thuringiensis. J. Econ. Entomol. 86: 226-233.
Worner, S.P. 1988. Ecoclimatic assessment of potential establishment of exotic pests. J. Econ. Entomol. 81: 973-983.
Wright, R.J. 1984. Evaluation of crop rotation for control of Colorado potato beetle (Coleoptera: Chrysomelidae) in commercial potato fields on Long Island. J. Econ. Entomol. 77: 1254-1259.
Zehnder, G.W. and G.K. Evanylo. 1988. Influence of Colorado potato beetle sample counts and plant defoliation on potato tuber production. Am. Potato J. 65: 725-736.
Zehnder, G.W. and J. Hough Goldstein. 1990. Colorado potato beetle (Coleoptera: Chrysomelidae) population development and effects on yield of potatoes with and without straw mulch. J. Econ. Entomol. 83: 1982-1987.
[Home]
Copyright 2008 Andrei Alyokhin
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}